

Активуючі рецептори природних кілерів
Цитотоксична дія природних кілерів залежить від динамічної рівноваги між інгібіторними та активуючими рецепторами. Для лізису клітини-мішені не достатньо самої лише відсунтності пригнічуючого сигналу, необхідна також наявність активуючого, який може надходити від одного із наступних рецепторів:
NKG2D
NCR
нектин і нектино-подібні рецептори
NKp80 та деяких інших.
Найкраще охарактеризований рецептор NKG2D, що розпізнає принаймні шість різних лігандів (три із них є трансмембранними білками (MICA, MICB та ULBP-4), ще три — гілкофосфатидилінозитол-заякореними (наприклад ULPB-4)). Ці ліганди відсутні на поверхні нормальних клітин організму, а з'являються у випадку генотоксичного або клітинного стресу, що виникає, наприклад, при зараженні вірусом або злоякісному переродженні.
Механізм цитотоксичної дії
Природні кілери можуть проявляти цитотоксичність одним із двох шляхів:
Більшість NK-клітин мають на своїй поврехні білок FasL, який при активації приєднується до рецептора смерті Fas на поверхні клітини-мішені і таким чином активує в ній апоптоз;
Природні кілери містять гранули заповнені білками перфоринами та гранзимами, при активації відбувається дегрануляція клітин. При цьому перфорини вбудовуються у мембрану мішені і утворюють канали, це саме по собі може викликатиосмотичний лізис клітини. Крім того, через ці канали у цитоплазму клітини можуть потрапляти протеази гранзими, які запускають каскад реацій активації апоптозу.
16.Схема антитілогенезу.
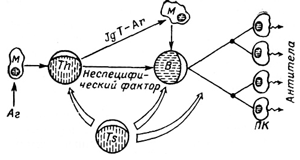
Условные обозначения:
Аг — антиген,
М — макрофаг,
Th — Т — хелпер (помощник),
Ts — Т — супрессор,
В — В — лимфоцит,
ПК — плазматические клетки.
Антителообразование — (син.: антителогенез нрк, биосинтез антител, синтез антител) образование специфических иммуноглобулинов, индуцированное антигеном; происходит главным образом в зрелых плазматических клетках, а также в плазмобластах и лимфобластах
Антитіло — білок-імуноглобулін, який виробляється в організмі у відповідь на антиген і здатний специфічно взаємодіяти з відповідним антигеном. Нині часто застосовують термін імуноглобулін. Ці терміни можна вважати синонімами.
Антитіло — фактор специфічного гуморального імунітету, молекула, яка розпізнає антиген. Антитіла є специфічними білками-імуноглобулінами. Їхня будова була визначена Г. Едельманом і Р. Портером, які вивчали так звані мієломні білки — продукти злоякісного розвитку певного клону лімфоцитів у хворих на мієлому. Інколи мієлома розвивається з однієї плазматичної клітини, тоді в організмі накопичується багато однакових молекул імуноглобуліну, які легко вивчати. Сьогодні з цією метою застосовують моноклональні антитіла.
Молекула будь-якого імуноглобуліну структурно організована за одним планом. Вона складається з двох типів ланцюгів — важких (Н — англ.heavy) та легких (L — англ. lіght). Ланцюги зв’язані дисульфідними містками. Легкі ланцюги мають молекулярну масу 20 кДа, містять 213–216 амінокислотних залишків і наполовину мають однакову первинну структуру незалежно від специфічності до антигену. Друга половина легких ланцюгів може мати багато варіантів, послідовність амінокислотних залишків у неї різна у антитіл до різних антигенів. Важкі ланцюги (молекулярна масса дорівнює 50 кДа, близько 500 амінокислотних залишків) складаються на 3/4 з константної та на 1/4 — з варіабельної частин.
Активний центр антитіла, який реагує з антигеном, називається паратопом. Він утворений варіабельними ділянками легких і важких ланцюгів. При розщепленні протеолітичними ферментами з молекули імуноглобуліну утворюються два або три фрагменти.
Імуноглобуліни за будовою та біологічними властивостями
поділяються на 5 класів.Імуноглобуліни кожного класу мають принципово однакову
організацію, вони складаються з одного або кількох мономерів. Мономер містить характерний для цього класу імуноглобулінів важкий ланцюг, а легкі ланцюги (їх може бути два варіанти — або обидва легкі -ланцюги, або -ланцюги) у всіх класів імуноглобулінів однакові, відрізняються лише по відношенню до антигену. Крім того, імуноглобуліни можуть містити додатковий J-ланцюг та додаткові компоненти. Усі молекули імуноглобулінів містять вуглеводи.
Синтез антитіл
Динаміка накопичення титрів антитіл після першого введення антигену (первинна імунна відповідь) характеризується такими закономірностями. Антитіла у певній кількості з’являються з 7-го дня, максимуму титр антитіл досягає на 10–15-й день, наприкінці місяця титри антитіл знижуються і лише незначно перевищують фонові.
При вторинній імунній відповіді титри антитіл збільшуються з другого дня, а зниження титрів антитіл відбувається значно повільніше, рівень титрів набагато вище.
Динаміка накопичення антитіл, які належать до різних класів імуноглобулінів, розрізняється при первинній і вторинній відповіді. При первинній відповіді можна побачити лише деяке відставання росту IgG порівняно з рівнем IgM. При вторинній відповіді титри IgM зростають практично так само, як і при первинній, тим часом як титри IgG зростають швидше за титри IgM, а далі довго утримуються на високому рівні. Тому менш ніж через місяць після вторинної стимуляції антигеном у крові виз начаються тільки IgG.
В синтезі антитіл розрізняють 2 фази: індуктивну та продуктивну.
Індуктивна фаза — це перші 24–72 год, коли відбувається засвоєння антигенної інформації, розмноження і диференціація клітин. Індуктивна фаза сприйнятлива до радіації та цитостатиків, наприклад, кортизону.
Продуктивна фаза може бути умовно поділена на клітинну (до 6 діб після введення антигену) та видільну (викид антитіл у кровообіг).