

Рис.
10-10. Эволюция генетических систем.
Каждый из кругов соответствует
определенному типу жизненного цикла.
Наиболее примитивные эукариоты
были, несомненно, гаплоидными на
протяжении большей части своего
жизненного цикла (круг, очерченный
одной линией, внизу слева). В этом случае
мейоз (обозначен четырьмя
жирными точками) происходит сразу
после оплодотворения (показано цветной
стрелкой). Другие жизненные циклы
отличаются от гаплоидного моментом
оплодотворения и протяженностью
диплоидной фазы (обозначена внешней
линией круга). Внутри кругов указаны
некоторые современные группы организмов,
характеризующиеся данным жизненным
циклом
ющие
питательные вещества из мертвых или
живых организмов. Грибы традиционно
относили к растениям, но сейчас уже не
осталось сомнений в том, что они являются
независимой эволюционной линией.
Кроме нитчатой структуры, грибы
практически не имеют ничего общего ни
с одной из групп, относимых к водорослям.
Их клеточные оболочки имеют хитиновый
матрикс; структуры, в которых формируются
споры, часто сложно устроены. Репродуктивные
циклы грибов, которые также могут быть
весьма непростыми, обычно включают как
половые, так и неполовые процессы. Этому
царству посвящена гл. 13.
Царство
Plantae
Это
царство объединяет моховидных (мхи,
печеночники, антоцеротовые) и девять
отделов сосудистых растений —
фотосинтезирующие организмы,
приспособленные к жизни на суше. Их
предками были специализированные
зеленые водоросли. Все растения
многоклеточные и состоят из эукариотических
клеток с вакуолями и целлюлозными
клеточными оболочками. Главный
способ питания — фотосинтез, хотя
некоторые виды стали гетеротрофными.
В ходе эволюции растений на суше
происходила их структурная диффе-
ренцировка с тенденциями к развитию
специализированных фотосинтезирующих,
заякоривающих и опорных органов (см.
рис. 10-7). У более сложно устроенных групп
такая организация привела к появлению
особых ассимиляционных, проводящих и
покровных тканей. Размножение растений
преимущественно половое с чередованием
гаплоидного и диплоидного поколений.
У более продвинутых представителей
царства гаплоидное поколение (гаметофит)
в ходе эволюции редуцировалось.
Моховидные рассматриваются в гл. 16, а
сосудистые растения в гл. 17, 18 и 29.
ПОЛОВОЕ
РАЗМНОЖЕНИЕ
ДНК
у бактерий не дуплицирована, как у
диплоидных эукариот; несмотря на
это, и у прокариот известно несколько
способов генетической рекомбинации.
Как более подробно описывается в гл.
11,
часть их генетического материала может
быть перенесена из одной клетки в
другую, но механизма, сравнимого с
мейозом, посредством
которого она затем регулярно передавалась
бы по наследству вместе с генетическим
материалом клетки-реципиента, здесь
нет.
Единственным
способом устойчивой передачи новой
генети- ' ческой информации у прокариот
является встраивание полученного
фрагмента в молекулу бактериальной
ДНК. Этот способ не точен и с трудом
воспроизводим.
Половое
размножение с регулярным чередованием
мей- оза и сингамии имеет высокое
селективное преимущество, поскольку
служит основным механизмом возникновения
и поддержания изменчивости природных
популяций эукариот. Изменчивость, как
мы узнаем из гл. 28, — важнейшая предпосылка
приспособляемости живых организмов к
окружающей среде в процессе эволюции.
Эволюция
диплоидии
Первые
эукариотические организмы были,
вероятно, гаплоидными и размножались
бесполым путем, однако с распространением
среди них полового размножения началась
эволюция диплоидии. По-видимому, она
возникла первоначально при объединении
двух гаплоидных клеток в диплоидную
зиготу. Такое эволюционное событие
скорее всего имело место неоднократно.
Можно полагать, что зигота затем
немедленно мейотически разделилась,
восстановив гаплоидное состояние (рис.
10-10 и 10-11,А).
У организмов с простым типом жизненного
цикла зигота является единственной
диплоидной клеткой.
«Случайно»
(а такой «случай» произошел в нескольких
разных эволюционных линиях) некоторые
из этих зигот поделились не мейотически,
а митотически, дав в результате начало
организму, состоящему из диплоидных
клеток, вступающих в мейоз
позднее. У животных подобный
отсроченный мейоз ведет
к образованию гамет — яйцеклеток и
спер- миев. Гаметы затем сливаются, что
непосредственно восстанавливает
диплоидное состояние (рис. 10-10
и 10-11,5). Таким образом, у животных гаметы
являются единственными гаплоидными
клетками.
У
растений мейоз приводит
к образованию спор, а не гамет. Споры —
это клетки, способные делиться
митотически, давая многоклеточный
гаплоидный организм; этим они отличаются
от гамет, которые могут развиваться
только после слияния друг с другом.
Многоклеточные гаплоидные особи,
чередующиеся с диплоидными формами,
характерны для растений, некоторых
бурых, красных и зеленых водорослей,
двух близкородственных родов
хитридиомицетов и других групп
протистов, не рассматриваемых в этой
книге.
152 Разд. IV. Разнообразие
Гл.
10. Классификация живых существ
Рис.
10-11. Схемы основных типов жизненных
циклов. Диплоидная фаза показана
снизу, а гаплоидная сверху от широкой
полосы. Четыре белые стрелки указывают
на продукты мейоза; одиночная белая
стрелка
— на
оплодотворенную яйцеклетку.
При
зиготическом мейозе зигота
делится мейотически, образуя четыре
гаплоидные клетки, делящиеся затем
митотически с образованием новых
гаплоидных клеток или многоклеточной
особи, которая в конечном итоге дает
гаметы за счет дифференцировки. Этот
тип жизненного цикла встречается
у хламидомонады и ряда других водорослей.
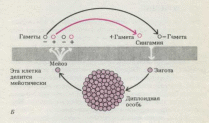
Б.
При гаметическом мейозе
диплоидная особь образует путем
мейоза гаплоидные гаметы, которые
сливаются, давая диплоидную зиготу. В
результате деления последней
возникает новый диплоидный организм.
Этот тип жизненного цикла характерен
для большинства животных, некоторых
протистов (оомицетов), а также для бурой
водоросли Fucus.
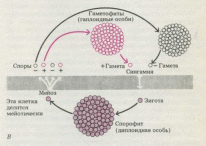
При
спорическом мейозе спорофит,
или диплоидная особь, в результате
мейоза образует гаплоидные споры, не
функционирующие как гаметы, а делящиеся
митотически. При этом возникают
многоклеточные гаплоидные организмы
(гаметофиты), формирующие в конечном
итоге гаметы, которые сливаются с
образованием диплоидной зиготы. Та
в свою очередь дает начало диплоидным
особям. Этот тип жизненного цикла
(чередование поколений) характерен
для растений и многих водорослей.
Сходный цикл наблюдается у хитридиомицета
Allomyces и еще одного близкого
к нему рода, а также у некоторых других
групп протистов, не включенных в эту
книгу
Такой
процесс известен как чередование
поколений
(рис. 10- 10 и 10-11, В).
Образующее гаметы гаплоидное поколение
у растений называется гаметофитом,
а диплоидное спорообразующее —
спорофитом.
Эта терминология используется также
в случае водорослей, а иногда и
применительно к другим группам.
У
ряда водорослей (большинство красных,
многих зеленых и отдельных бурых)
диплоидная и гаплоидная формы внешне
одинаковы. Такие типы жизненных циклов
называют изоморфным
чередованием поколений (рис. 10-10).
Имеются
и другие жизненные циклы, в которых
диплоидные и гаплоидные формы не
идентичны. В ходе эволюции
некоторых
групп происходили мутации, которые
проявлялись только в одном поколении,
хотя соответствующие аллели были,
очевидно, представлены и у диплоидных,
и у гаплоидных особей. В жизненных
циклах такого типа гаметофит и спорофит
стали заметно отличаться друг от друга
— возникло гетероморфное
чередование поколений, характерное
для растений и некоторых бурых водорослей
(рис. 10-10).
У
моховидных (мхов, печеночников,
антоцеротовых) гаметофит доминирует;
он независим в своем питании и обычно
крупнее спорофита, который может быть
структурно более сложным. У сосудистых
растений, напротив, доминирует спорофит,
который намного крупнее и сложнее
153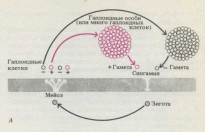
154
Разд. IV. Разнообразие
гаметофита,
почти у всех групп зависимого в своем
питании от спорофита.
Как
уже упоминалось раньше, диплоидия
позволяет накапливать больше
наследственной информации и гораздо
тоньше регулировать проявление
генетического материала организма в
ходе его развития. Возможно, в этом
причина того, что спорофит у сосудистых
растений — крупное, сложное,
независимое в отношении питания
поколение. Одна из наиболее четких
эволюционных тенденций у этой группы,
преобладающей в большинстве наземных
местообитаний, — усиление доминирования
спорофита и подавление гаметофита.
У цветковых растений женский гаметофит
— микроскопическое тельце всего из
семи клеток, а мужской — только из трех
клеток. Оба они полностью зависят от
спорофита в смысле питания.
ЗАКЛЮЧЕНИЕ
Биологи
разработали методы наименования и
классификации живого, позволяющие
очень точно обозначать организмы,
что является важнейшим фактором научного
обмена информацией. Классификация дает
возможность показать взаимосвязи
данной формы с другими живыми существами.
Организмы
имеют научное, состоящее из двух слов
(бинарное) название. Первое слово —
название рода, а второе — видовой
эпитет; вместе они дают полное название
вида. Виды иногда подразделяются на
подвиды или разновидности. Роды
объединяются в семейства, семейства в
порядки, порядки в классы, классы в
отделы, отделы в царства. Царство —
высшая единица классификации живого.
В
этой книге живые организмы сгруппированы
в пять царств, первое из которых
объединяет прокариот, остальные —
эукариот: (1)
Мопега (бактерии, или прокариоты); (2)
Protista
(простейшие, эукариотические водоросли,
слизевики и водные плесени); (5) Animalia
(многоклеточные нефотосинтезирующие
организмы); (4)
Fungi
(грибы) и (5) Plantae
— моховидные и сосудистые растения —
фотосинтезирующие организмы,
населяющие сушу и более сложно устроенные,
чем водоросли.
Митохондрии,
характерные для всех эукариот, вероятно,
возникли из аэробных (нуждающихся в
кислороде) прокариот, сходных с
пурпурными несерными бактериями, а
хлоропласти, по-видимому,
произошли по крайней мере от трех
различных групп фотосинтезирующих
аэробных бактерий. Поэтому наличие
хлоропластов у разных групп эукариот
не означает прямых родственных связей
между ними. Много- клеточность возникла
неоднократно и независимо у разных
групп протистов. Некоторые из
многоклеточных эволюционных линий,
включая красные, бурые и зеленые
водоросли, отнесены в используемой
нами системе классификации, признающей
пять царств, к протистам, тогда как три
основные многоклеточные линии —
животные, растения и грибы — считаются
самостоятельными царствами.
В
эволюции организмов диплоидия появилась
вслед за процессом полового размножения.
У примитивных эукариот и всех грибов
зигота, возникающая при сингамии, сразу
же делится мейотически. От древних
жизненных циклов такого
рода
в нескольких эволюционных линиях
независимо произошли более сложные,
включающие диплоидные фазы с митотически
делящейся зиготой. Если гаплоидные
клетки, появляющиеся при мейозе,
сразу
же функционируют как гаметы, речь идет
о жизненном цикле, типичном для животных
и некоторых групп протистов. Если же
они делятся митотически, как у водорослей,
всех растений и двух родов хитридиомицетов,
их считают спорами. Диплоидное поколение,
дающее споры, называется спорофитом,
а гаплоидное, развивающееся из спор в
результате митозов, — гаметофи- том. В
конечном итоге за счет митоза оно дает
начало гаметам. Если гаметофит и
спорофит в жизненном цикле приблизительно
одинаковы по размеру и сложности
строения, то цикл называется изоморфным;
если же они по этим признакам сильно
различаются — гетероморфным.
РЕКОМЕНДУЕМАЯ
ЛИТЕРАТУРА
Altschul,
Siri von Reis:
“Exploring the Herbarium”, Scientific American, May 1977.
pages 96—104.
О
том, какие ценные сведения можно
почерпнуть из гербарных этикеток об
использовании малоизвестных растений
как источника пищи, лекарств или других
полезных продуктов.
Bold,
Harold G., Alexopoulos С.
S., Delevoryas Т.:
Morphology of plants and fungi, 4th ed., Harper & Row,
Publishers, Inc., New York. 1980.
Хорошо
иллюстрированная, содержательная книга
о разнообразии растений, водорослей
и грибов.
Comer
Е.
J. Н.:
The life of plants, Mentor Books, New American Library, Inc., New
York, 1968.
Известный
ботаник поэтично описывает эволюцию
растений, изменения их структур и
функций при столкновении с особенностями
новой среды во время расселения по
суше.
Jones
S. В.,
Luchsinger Arlene Е.:
Plant systematics, McGraw-Hill Book Company, New York, 1979.
Полный
хороший курс практической систематики.
Margulis
Lynn:
Symbiosis in cell evolution, W. H. Freeman and Company, New
York, 1981.
Увлекательное
повествование о происхождении
эукариотических клеток в результате
установления нескольких симбиозов.
Margulis
Lynn, Schwartz Karlene V.:
Five kingdoms. An illustrated guide to the phyla of life on Earth,
W. H. Freeman and Company, New York, 1982.
Прекрасное
сжатое описание разнообразия жизни на
Земле.
Parker,
Sybil P.
(Ed.): Synopsis and classification of living organisms, 2
vols., McGraw-Hill Book Company, Inc., New York, 1982.
Энциклопедический
авторитетный обзор основных групп
растений, животных и микроорганизмов.
Scagel,
Robert F.
et al. Nonvascular plants: an evolutionary approach, Wadsworth
Publishing Co., Inc., Belmont, Calif., 1982.
Исчерпывающий,
тщательно иллюстрированный обзор
разнообразия моховидных, грибов,
бактерий и протистов, рассматриваемых
в нашей книге.