

Сенсорные оценки - ощущения и восприятия
Ощущения - это основные данные органов чувств, сырой материал, из которого извлекается знание. Красное и синее - это примеры цветовых ощущений. Восприятие - это процесс как интерпретации сенсорной информации в свете опыта, так и бессознательного заключения. Например, глядя на какой-нибудь пейзаж, мы используем цвет как руководство для суждения о расстоянии, на каком находятся холмы и другие объекты на местности. Это хорошо знают художники, которые обычно пользуются разными оттенками синего, чтобы создать впечатление расстояния.
Психологи изучают восприятие у людей, главным образом в лаборатории, и обычно испытуемые должны давать словесный ответ в различных условиях восприятия. Исследованию ощущения и восприятия у животных мешает то, что они не могут разговаривать, и трудно представить, как экспериментальные ситуации интерпретируются животными. Но многие из методов, разработанных исследователями психологии человека, нашли применение в изучении поведения животных.
Фехнер (1801-1887) полагал, что ощущения нельзя измерить непосредственно, так как они представляют собой полностью личный опыт. Можно измерить величину физического стимула, а человеку предложить оценить величину полученного ощущения. Проблема состоит в том, что, если два человека дадут разные количественные оценки одинаковым стимулам, мы не можем сказать, получили ли они разные ощущения или же просто оценили по-разному их величину. Фехнер понимал, что, хотя ощущения нельзя сравнить с физическими стимулами, они сравнимы между собой. Человек может сравнить два ощущения и сказать, одинаковые они или разные. Один из методов изучения этой способности - постепенное изменение интенсивности стимула, предъявляемого испытуемому, пока он не скажет, что заметил разницу. Эта разница между измененной и исходной интенсивностью, устанавливаемая в результате неоднократных повторений опыта, названа едва заметной разницей. Ее физическая величина может быть измерена в широком диапазоне интенсивностей физического стимула. Например, испытуемому можно предложить несколько раз сравнить две массы: одну - стандартную, скажем 100 г, а вторую - из набора для сравнения. Испытуемый должен ответить, была ли вторая масса больше первой, такой же или меньше ее. На основании повторных тестов такого типа экспериментатор может определить, во-первых, интервал неопределенности, в пределах которого испытуемый не уверен, отличается ли предлагаемая масса от стандартной, а во-вторых, разностный (дифференциальный) порог, то есть величину, на которую надо изменить интенсивность стимула, чтобы вызвать другое ощущение.
В 1834 г. немецкий физиолог Вебер высказал мысль, что разностный порог представляет собой постоянную долю стандартного стимула. Иными словами, если изменение массы от 100 до 110 г (на 10%) составляло едва заметную разницу, то разница между 10 и 11 г, т.е. то же изменение на 10%, должно быть различимо точно так же. Другими словами, если I - это значение стандартного стимула, а DI - прирост его интенсивности, необходимый для возникновения едва заметной разницы, то отношение ∆I / I = const. Это выражение известно как закон Вебера, а дробь DI /I -это отношение Вебера.
Фехнер считал, что закон Вебера служит ключом к измерению субъективного опыта. Он и его сотрудники провели множество опытов для проверки этого закона. В общем, он правилен для нормальных пределов интенсивности стимула, но может нарушаться в краевых областях этого диапазона интенсивности. Фехнер особенно интересовался соотношением между физической интенсивностью стимула и субъективной величиной вызываемого им ощущения. Он предложил следующую формулу, известную под названием закона Фехнера:
S = k log I,
где S - субъективная величина, I - интенсивность стимула, а k - константа. Интересным следствием такого подхода является то, что разным сенсорным модельностям присущи разные отношения Вебера, как показано в табл. 12.2. Чем меньше отношение, тем тоньше ощущение. Так, среди испытанных видов чувствительности у человека различительная способность зрения наибольшая, а вкуса наименьшая.
Таблица 12.2. Типичные значения отношения Вебера для разных видов чувствительности

В целом это теоретическое и экспериментальное направление названо психофизикой. Законам Вебера и Фехнера предложены разные альтернативы, и некоторые исследователи считают, что степенной закон дает более точную интерпретацию данных. Но против классического психофизического подхода были выдвинуты и более основательные возражения.
Классический психофизический эксперимент требует, чтобы испытуемый отвечал по принципу «да-нет». Интерпретация результатов предполагает, что существует совершенно определенный критерий, для того чтобы сказать: «Да, я замечаю разницу» или «Нет, не замечаю», т. е. считается, что испытуемый не дает неопределенных ответов. Ранние исследователи знали, что состояние человека может изменять даваемые им в опыте оценки. Они пытались устранить это неудобство, работая только с тщательно тренированными испытуемыми или вводя время от времени обманные пробы и учитывая возможность ложных ответов. Коренное улучшение было получено введением методов, заимствованных из теории обнаружения сигналов.
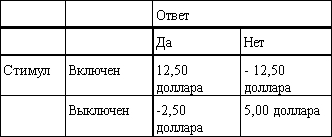
Эта теория не использует четкого критерия «да - нет». Она считает, что ответ «да» или «нет» испытуемого зависит от действия стимула с учетом существующей изменчивости (шума) в уровне данного раздражения от ожидания наблюдателя и от потенциальных последствий его решения. Предположим, например, что испытуемому предложено ответить, громче ли подаваемый сигнал, чем стандартный. Четыре возможных результата приведены в табл. 12.3. Предположим также, что мы условились вознаграждать испытуемого суммой в 25 центов за каждую правильно обнаруженную разницу в громкости (попадание) и штрафовать его на 25 центов за промах. Мы платим также по 10 центов за каждое правильно установленное отсутствие разницы (правильное отрицание) и штрафуем на 5 центов за указание разницы, когда ее не было (ложная тревога). Теперь результаты ответов испытуемого можно выразить в виде платежной ведомости (табл. 12.4). Допустим, испытуемому предложено 100 тестов, в одной половине которых имеется истинная разница в громкости, а в другой половине разницы нет. Неизменно говоря «да», он заработал бы 12 долларов 50 центов за 50 правильных ответов и был бы оштрафован на 2 доллара 50 центов за 50 ошибок. Неизменно говоря «нет», испытуемый заработал бы 5 долларов за 50 правильных ответов и был бы оштрафован на 12 долларов 50 центов за 50 ошибок. Таким образом, отвечая все время положительно, испытуемый заработал бы 10 долларов, а отвечая все время отрицательно, потерял бы 7 долларов 50 центов. Неудивительно, что при таком расчете испытуемый был бы настроен отвечать «да».
Таблица 12.3. Четыре возможных результата опыта по обнаружению стимула

Таблица 12.4. Платежная ведомость. Доллары, заработанные и потерянные при ответах «да» и «нет», когда стимул включен или выключен в 100 пробах
Другая форма настроенности создается ожиданиями испытуемого. Так, например, Линкер и др. обнаружили, что в тестах тенденция давать положительный ответ отчасти зависела от того, как часто предъявлялся стимул. Если он предъявлялся в 90% случаев, авторы получали большой процент «попаданий» и ложных тревог. Когда же этот стимул предъявлялся только в 10% случаев, правильных положительных ответов и ложных тревог оказывалось мало.
Согласно теории обнаружения сигналов, не существует нулевых стимулов, т. е. испытуемый знает, что сенсорные события засорены фоновым шумом. Поэтому задача состоит не в том, чтобы просто сказать, предъявлен ли стимул, а в том, чтобы отличить его сочетание с шумом от одного шума. Критерий решения устанавливается тенденцией испытуемого к определенному ответу, которая в свою очередь определяется системой вознаграждения. Так, при одном способе проверки этой теории условия стимуляции оставляют постоянными, а меняют систему оплаты. При другом способе сохраняют тенденцию отвечать определенным образом, а меняют свойства стимуляции. Результат такого рода опыта по определению тенденции к определенному ответу получается очень достоверный. У более консервативного испытуемого понижается доля как ложных тревог, так и правильных попаданий, и наоборот. При усилении стимула число ложных тревог снижается, а число попаданий растет.
Психофизическими методами изучались многие стороны поведения животных. Первым исследованием такого рода, по-видимому, была работа Блафа (Blough, 1955), который изучал темновую адаптацию у голубей. Этот автор научил голубей клевать два ключа, расположенных непосредственно под освещенным окошечком. Свет проникал в аппарат только от этого источника. Птицы были научены клевать ключ А, когда видели стимул, и ключ Б, когда не видели его. Клевание ключа А автоматически приводило к потускнению стимула, а клевание ключа Б повышало его яркость. Обученную птицу тестировали следующим образом.
Сначала стимульное окошечко ярко освещено, и птица клюет только ключ А. Каждый удар клювом снижает яркость стимула, но птица продолжает клевать этот ключ, пока стимул не станет таким слабым, что будет ниже абсолютного порога для голубя. В этот момент птица начинает клевать ключ Б, из-за чего стимул становится ярче. Когда он снова становится видимым птице, она возвращается к ключу А. В течение часа птица попеременно клюет ключи А и Б, и яркость стимула колеблется в области абсолютного порога птицы. Эта процедура позволяет проследить кривую темновой адаптации голубя. Сначала птица адаптирована к свету, но по мере потускнения стимула начинает адаптироваться к темноте. Иными словами, ее порог постепенно меняется. Перегиб этой кривой, соответствующий переходу от колбочкового к палочковому зрению.
Фильтрация стимула
Анатомические и физиологические исследования органов чувств и связанных с ними отделов нервной системы могут дать ценную информацию о сенсорных способностях животного. Однако сами по себе они не позволяют сделать окончательных выводов о том, что животное воспринимает и чего не воспринимает, поэтому обычно желательно поведенческое подтверждение. Кроме того, демонстрация того, что ЦНС получает определенную сенсорную информацию, ничего не говорит нам о том, как эта информация используется.
Потенциально животному доступно гораздо больше информации, чем оно в состоянии зарегистрировать и на которую способно ответить. Каким-то путем животное должно производить отбор, реагируя только те явления, которые ему важны, и игнорируя остальные. Этот феномен называют фильтрацией стимула. Смысл его в том, что на разных стадиях причинной цепи между стимулом и реакцией некоторые стимулы отфильтровываются и не влияют на поведение животного.
В известном смысле некоторая степень фильтрации обусловлена ограниченными возможностями органов чувств. Так, например, ухо человека не реагирует на звуки частотой выше 20 кГц. Глаз человека отфильтровывает инфракрасные и ультрафиолетовые части спектра, хотя некоторые животные их различают. Интересный пример приведен в работе по древесной лягушке. Ее видовое латинское название происходит от особого издаваемого самцом звука «ко-кви», который служит для привлечения самок и отпугивания самцов. Барабанная перепонка у самцов и самок настроена по-разному. Самцы слышат только звук «ко», а самки - звук «кви». Точно так же рецепторы насекомых часто высокоспециализированны и реагируют только на узкий диапазон стимуляции.
Когда стимул обнаружен, он может быть автоматически классифицирован таким образом, что все его посторонние особенности отфильтровываются. Так, например, Летвин и др. показали, что у лягушки фоторецепторы сетчатки соединены между собой, образуя рецептивное поле. Некоторые из них, называемые «детекторами жуков», различают главным образом мелкие темные движущиеся предметы. В поисках пищи лягушки реагируют на такие объекты больше, чем на другие стимулы.
Избирательная реактивность широко распространена у животных. Реагируя в определенной ситуации, животное использует только часть потенциально доступной информации. Так, например, Лэк наблюдал, что самцы зарянок часто нападают на других красногрудых зарянок, которые вторгаются на их территорию. Они нападут также на чучело зарянки, помещенное на эту территорию, но только если у него красная грудь. По-видимому, красная грудь служит мощным стимулом для выделения нарушителя границ. Лэк показал, что хозяин территории энергично нападает на пучок красных перьев, как будто это зарянка-нарушитель. Нет сомнения, что зарянка способна отличить пищу от пучка перьев, но в ситуации защиты территориальных границ она как бы слепнет в отношении всех прочих признаков объекта, кроме красной груди. Такого типа стимул называется сигнальным раздражителем.
Сигнальные раздражители могут меняться с изменением внутреннего состояния животного. Так, серебристые чайки крадут и поедают яйца других чаек. Чайка-грабитель различает яйца по их форме. Но для насиживающей чайки при возвращении выкатившихся яиц в гнездо важнее всего их величина и окраска, а форма имеет сравнительно малое значение. Такие птицы, в частности, готовы затаскивать в гнездо круглые или цилиндрические предметы того же размера и окраски, что и настоящее яйцо. Однако, как только чайка садится на яйца, их форма снова приобретает значение и если отличается от округлой, птица отказывается от насиживания. Таким образом, яйца обладают тремя разными наборами сигнальных раздражителей соответствующими трем разным видам поведения.
Другой интересный пример сигнальных раздражителей содержится в работе Г. Бургхардта с сотрудниками по пищевому поведению подвязковых змей и ужей, которые живут в США в реках и прудах и питаются мелкими рыбешками и червями. Они обнаруживают свою добычу по вкусу и запаху и обладают хеморецепторами, находящимися в парных ямках на нёбе, так называемым «якобсоновым органом». Змея высовывает язык, слизывает химические вещества с добычи и затем вставляет кончик языка во вкусовые ямки. Бургхардт предлагал разным видам подвязковых змей ватные тампоны, пропитанные экстрактами из рыб, лягушек, саламандр и червей. Он обнаружил, что разные виды оказывали большее предпочтение тому типу добычи, которым они обычно питаются в естественных условиях. Эта тенденция проявлялась у ни разу не питавшихся новорожденных змей, причем ее нельзя было изменить, меняя пищу матери или насильно кормя молодых змей искусственной пищей. Однако предпочтения изменяются, когда змеи переключаются на другую добычу. Так, виды, в естественных условиях предпочитавшие гольянов, начинают предпочитать серебряных карасей, привыкнув питаться этими рыбами. По-видимому, химические свойства определенной добычи действуют как сигнальные раздражители в отношении которых змея обладает генетически обусловленной склонностью, однако эта склонность может изменяться под влиянием приобретенного опыта.
Хотя некоторые сигнальные раздражители могут быть результатом периферической фильтрации, совершенно очевидно, что большая часть таких стимулов определяется процессами в ЦНС, какой-то центральный фильтрующий механизм определяет способность многих видов предпочтительно реагировать на сигнальные раздражители.
В этологических исследованиях также привлек к себе внимание особый временный тип фильтрации стимулов, отличный от сравнительно постоянного опознания сигнальных раздражителей. Это понятие поискового образа, впервые предложенное Я.Юкскюлем. Нам всем знаком такой феномен восприятия, когда вдруг увидишь то, чего раньше не замечал. Когда мы смотрим на фотографию замаскированных насекомых, то сначала можем не различать ни одного из них, потом вдруг увидим одно, за ним другое и т. д. После этого нам кажется, что найти их на фотографии совсем нетрудно. У нас сформировался поисковый образ насекомого.
Во многих работах показано наличие таких поисковых образов у животных. Так, например, Кроуз научил черных ворон искать корм, спрятанный под раковинами мидий разного цвета, разбросанными на большом расстоянии друг от друга по земле. Вороны должны были переворачивать каждую раковину, чтобы увидеть, не лежит ли под ней корм. В течение некоторого времени они предпочтительно сосредоточивали поиск на одном цвете и игнорировали раковины других цветов, хотя у них и был опыт обнаружения корма под раковинами всех типов. Одного образца, предъявленного экспериментатором, иногда было достаточно, чтобы вызвать поиск раковин определенного цвета. Вороны вели себя так, как будто обладали поисковым образом для определенной окраски раковины, который сохранялся некоторое время, но легко мог переключиться на другой цвет.
Самое тщательное исследование поисковых образов проведено Мэриан Доукинс у цыплят в лабораторных условиях. В течение трех недель цыплят кормили окрашенными в оранжевый или зеленый цвет рисовыми зернами на белом фоне. Затем тестировали способность цыплят обнаруживать зерна на фоне другого цвета и на фоне того же цвета. Доукинс обнаружила, что, хотя сначала цыплята не различали замаскированные зерна, постепенно по мере приобретения опыта наступало заметное улучшение их поиска. Выяснилось также, что опыт поедания заметных зерен понижал способность цыплят видеть замаскированный корм. Поэтому способность обнаруживать его, очевидно, не может объясняться просто приобретением опыта.
В следующих экспериментах Доукинс тестировала цыплят, которые уже питались определенным исходным видом зерен. Она установила, что, когда исходные зерна были заметны, цыплята больше реагировали на тестовые зерна, выделяющиеся по цвету. Когда же исходные зерна были замаскированы, цыплята больше реагировали на тестовые зерна, различимые не по цветовым признакам, а по текстуре и форме. Эти данные можно толковать как избирательное внимание. Так, когда исходные зерна были заметны благодаря их цвету, цыплята обращали внимание на цвет, и, следовательно, затем им было легче обнаруживать зерна по цвету. Когда же исходные зерна были замаскированы, т. е. окрашены точно так же, как фон, цыплята обращали внимание на нецветовые признаки. В результате им было легче обнаруживать тестовые зерна по нецветовым признакам.
Животное способно различать два стимула, но обычно лишь после того, как его научат этому. Такое научение называется дискриминационным, или различительным. Подходящими стимулами для исследования такого научения у крыс являются черный и белый прямоугольники. Обычно их предъявляют на сером фоне, и опыт должен содержать все рассматриваемые ниже обычные контроли. Не ограничивая животное одним признаком, важнейшим при исследовании сенсорных способностей, можно изучать, как наличие двух путей решения задачи влияет на процесс научения. Так, прямоугольники могут различаться величиной или ориентацией.
Рассмотрим случай различения по яркости и ориентации, когда черный прямоугольник предъявляется в вертикальном положении, а белый - в горизонтальном. Теперь мы должны решить, какой стимул будет вознаграждаться, а какой не будет. Здесь мы наталкиваемся на проблему, как не спутать предпочтения крысы с ее способностями к научению. Они предпочитают черное белому. При вознаграждении черного стимула крысе будет легче решить задачу, чем при вознаграждении белого. Та же проблема встает и в отношении ориентационного признака, так что 20 крыс поделили на 4 группы.
Таблица 5. План опыта по различению
Группа |
Число крыс |
Ориентация стимула |
Цвет стимула |
1 |
5 |
Г |
Ч |
2 |
5 |
В |
Ч |
3 |
5 |
Г |
Б |
4 |
5 |
В |
Б |
Ч -черный; Б -белый; Г -горизонтальный; В - вертикальный. |
|||
Сазерленд и Макинтош показали, что при решении задач такого типа животные должны научиться двум вещам: 1) на какие свойства стимулов обращать внимание и 2) какое из двух проявлений этого свойства вознаграждается. Например, крыса, получившая награду за то, что обратила внимание на яркость стимула, скорее будет и в дальнейшем обращать внимание на яркость. Крысы, научившиеся реагировать на определенную яркость или ориентацию или на оба этих свойства, успешно научались решать описанную выше задачу.
Чтобы определить, на какие свойства стимулов крысы в самом деле научились обращать внимание, проводятся невознаграждаемые тесты на перенос. При них крысам предъявляют стимулы, различные по ориентации или яркости, но не по обоим этим свойствам. Так, в половине проб стимулы представляют собой черные или белые прямоугольники, различающиеся только по ориентации, а во второй половине горизонтальные или вертикальные прямоугольники, различающиеся только по яркости. Крысы, научившиеся обращать внимание лишь на ориентацию, способны решить только первую задачу, а те, которые научились обращать внимание только на яркость, могут решить вторую задачу, но не могут — первую.
Экологическая обусловленность чувств животных.
Многие различия между животными объясняются не столько их эволюционной историей, сколько экологическими условиями.
Животное, глаза которого приспособлены к яркому свету, будет обладать хорошей остротой зрения, восприятием цвета и движения. Такие типично дневные глаза относительно нечувствительны к низким уровням освещенности. Животные, адаптированные к сумеречному освещению, обладают более высокой чувствительностью, но в ущерб цветовому и детальному предметному зрению.
Видеть при слабом освещении нужно животным, ведущим ночной образ жизни, обитающим глубоко в воде и в пещерах. Эти условия не совсем сравнимы, потому что спектр приходящего света смещен в воде. Нет данных о том, что ночные животные обладают повышенной чувствительностью к длинным волнам. Однако некоторые из них владеют приспособлениями, повышающими чувствительность к свету. Глаза с широким зрачком и большим хрусталиком улавливают больше света, чем маленькие глаза. Глазами первого типа обладают, например, опоссум, домовая мышь и рысь. У других ночных животных, например сов и галаго, череп сужен с боков, что привело к цилиндрическому удлинению глаза. Цилиндрические глаза встречаются также у некоторых глубоководных рыб. Многие глубоководные животные специально адаптированы к господствующим условиям освещения. Максимумы поглощения у зрительных пигментов глубоководных рыб совпадают с длиной волны, максимально пропускаемой водой. У глубоко ныряющего кита северного плавуна зрительные пигменты сильнее всего поглощают более короткие волны, чем у неглубоко ныряющею серого кита. Точно так же глаза глубоководных морских ракообразных содержат зрительные пигменты с максимумами поглощения в более коротковолновой области, чем у мелководных ракообразных.
В дополнение к свойствам зрительных пигментов сумеречное зрение усиливают также другие специальные приспособления. Как правило, у ночных животных палочек больше, чем колбочек, а у некоторых из них, например у кошачьей акулы и галаго, мало или совсем нет. У многих ночных позвоночных близ фоторецепторов расположены отражающие свет образования, благодаря которым глаза "светятся", когда на них падает луч света. Свет, который проходит через фоторецептор не поглотившись, отражается обратно, что повышает вероятность поглощения.
Существует грань, за которой условия для зрения так тяжелы, что от глаз как от главного органа чувств приходится отказаться. У большинства животных, обитающих в пещерах, в глубине моря или же в мутной воде, глаза рудиментарные вследствие их регрессии. Например, пещерные земноводные обладают глазами на личиночной стадии, но лишены их во взрослом состоянии. Если этих животных воспитывать на свету, то у взрослых особей развиваются нормальные глаза. У молоди слепой пещерной рыбы имеются глаза, но у взрослых животных они дегенерируют. Садоглу обнаружил, что дегенеративное состояние глаз у пещерного вида определяется генами.
Среди млекопитающих кроты и летучие мыши являются самыми известными примерами дегенерации зрения. Очень маленькие глазки кротов у некоторых видов покрыты кожей. У плотоядных рукокрылых зрение развито хорошо, но у ночных летучих мышей, особенно у тех видов, которые ловят насекомых на лету, зрение очень слабое.
Заменители зрения
Животные с очень слабым зрением, живущие при сумеречном свете, должны рассчитывать на другие органы чувств. Так, у придонных кошачьих сомов и ошибневых имеются разные сенсорные усики, которыми они ощупывают субстрат. Эти усики снабжены многочисленными осязательными рецепторами и хеморецепторами. Однако такие органы чувств не могут быть источником информации о величине и положении предметов в окружающей среде. Рыбы с нейромастами и органами боковой линии, чувствительными к вибрации, могут обнаруживать движущиеся предметы и получать некоторую информацию о неподвижных предметах по отраженным от них движениям воды. Но наилучшей заменой зрения животным служат электромагнитная чувствительность и особые слуховые приспособления.
Некоторые виды рыб используют электрическую чувствительность при обычной ориентации и коммуникации. Чувствительность к магнитным полям тоже обнаружена у ряда животных. Так, некоторые бактерии ориентируются к северному магнитному полюсу и реагируют на магнит в лабораторных условиях. С помощью электронного микроскопа у таких бактерий были обнаружены цепочечные структуры, содержащие кристаллы магнетита, которые также найдены в брюшке медоносных пчел и в сетчатке голубей. В Северном полушарии бактерии следуют наклонению магнитного поля Земли, и оно направляет их вниз, в анаэробный ил, их естественную среду обитания. В Южном полушарии у бактерий обратная полярность.
Рыбы используют электричество тремя разными способами.
1) Так называемые "сильноэлектрические" рыбы, такие, как электрический скат и электрический угорь, производят электрические разряды, способные оглушить жертву, но лишены электрической чувствительности.
2) Электрочувствительные рыбы, например Scyliorhinus и некоторые другие акулы, не производят электричества. Однако кошачьи акулы способны обнаружить даже зарывшуюся в песок добычу по локальному искажению электрического поля Земли. Для этого служат особые органы чувств - так называемые ампулы Лоренцини, широко распространенные по всей поверхности тела, особенно вблизи головы.
3) Так называемые "слабоэлектрические" рыбы генерируют свои электрические поля и чувствительны к электрическим изменениям среды. Обычно это ночные рыбы, обитающие в мутной воде. У них два типа электрочувствительных рецепторов: ампульные, которые отвечают на медленно изменяющиеся электрические поля, и клубневые, реагирующие только на быстрые их изменения. У некоторых видов один тип рецепторов, у других - оба. Эти рыбы генерируют слабые электрические поля посредством электрических органов, которые представляют собой видоизмененные мышцы или аксоны. Электрические разряды, как правило, испускаются с частотой до 300 импульсов в секунду. Некоторые рыбы способны менять частоту импульса. Когда одна рыба испытывает электрическое воздействие другой, она может изменить частоту своих импульсов, чтобы снизить это вмешательство. Электрорецепторы служат также для локализации предметов в окружающей воде по искажениям, которые эти предметы вызывают в электрическом поле. Некоторые рыбы, например Gymnarchus, отличают хорошие проводники от плохих, например металлический стержень от пластикового.
Слух заменяет зрение многим видам, и у некоторых из них образовались весьма интересные и специализированные добавления к нормальному слуху. Все эти адаптации способствуют точной локализации источника звука. Сравнение сигналов, приходящих в оба уха, служит главным способом локализации этого источника у позвоночных. Человек с одним ухом может, поворачивая голову, искать направление максимальной силы сигнала, потому что голова создает определенную звуковую тень. Два уха делают возможным одновременное сравнение, что позволяет локализовать источник гораздо быстрее и точнее. Если уши достаточно далеко отстоят друг от друга, то создается разница во времени прихода, в фазе звуковых волн, имеющих определенное направление, и их интенсивности. Таким образом, мелкие животные сравнивают только интенсивность звуков, а люди пользуются и монауральными, и бинауральными способами, и долгое время считалось, что они являются видом с наилучшей способностью к обнаружению источника звука. Но благодаря работе Пэйна, по слуховым способностям сова сипуха далеко превосходит человека. Она может локализовать и поймать свободно движущуюся мышь в полной темноте. Она даже способна определить направление движения животного и благодаря этому располагает свои когти вдоль длинной оси его тела. Сипуха пользуется низкочастотными компонентами звуков для локализации их источника в горизонтальной плоскости, а высокочастотными - для определения его положения по вертикали. Она не смешивает эти два типа информации, даже несмотря на то, что оба они основаны на сравнении звуков, приходящих в оба уха.
Самой совершенной заменой зрения является эхолокация, при которой животное испускает высокочастотные сигналы и обнаруживает предметы по возникающему от них эху.
Простые формы эхолокации встречаются у землероек, жирных козодоев и гималайских саланганов, которые ночуют и гнездятся в пещерах. Более совершенными ее формами обладают дельфины и другие морские млекопитающие, но своей вершины она достигает у летучих мышей.
В процессе эхолокации летучие мыши испускают залпы ультразвуковых импульсов малой длительности и высокой частоты. Такие короткие импульсы позволяют точно определять время образования эха, а значит, и расстояние до образующего его предмета. Звуки, производимые другими животными и ветром, обычно бывают низкочастотными, поэтому маловероятно, чтобы на ультразвуковые сигналы летучих мышей накладывались помехи. Однако, искусственные звуки частотой выше 20 кГц дезориентируют полет летучих мышей. Другое преимущество высоких частот состоит в возможности точной фокусировки, что делает возможным распознавание мелких объектов.
У летучих мышей много также специальных приспособлений, позволяющих им определять время и локализовать место возникновения эха от их ультразвуковых сигналов. У большинства рукокрылых, хватающих насекомых на лету, большие наружные уши, форма которых повышает дирекциональную чувствительность.
Зрительное опознание жертвы и хищника
Большинство хищников встречается с множеством разных видов, которые служат им добычей и которых необходимо отличать от других организмов. При этом чаще всего используются три признака - величина, движение и форма.
Когда хищники должны выбирать между особями-жертвами, отличающимися только размерами тела, они обычно хватают самую крупную. Эта стратегия наиболее эффективна в отношении затрат энергии. Однако есть предел, за которым стимул больше не воспринимается как добыча. Например, когда обыкновенной жабе предъявляют как "добычу" объекты разной величины, она реагирует положительно на экземпляры, размеры которых находятся в определенных пределах, но активно избегает более крупные стимулы. Чтобы выбрать добычу определенной величины, жабе нужно оценить абсолютную величину видимого объекта, учитывая и его размеры на сетчатке, и расстояние до него. Жабы-повитухи в процессе развития постепенно научаются выбирать добычу постоянного размера.
В некоторых случаях наблюдается склонность к более крупной добыче, но не из-за активного предпочтения, а потому, что она лучше различима. Так, радужные форели чаще ловят крупных ракообразных, чем мелких, потому что крупные видны на большем расстоянии.
Для того чтобы узнать добычу, некоторым видам, например лягушкам и жабам, нужно, чтобы она двигалась. Обыкновенная каракатица в норме нападает только на движущихся креветок. Но если у нее отнять только что пойманную и парализованную ею креветку, то это головоногое немедленно снова нападет на нее даже на неподвижную. Некоторые хищники предпочитают добычу, которая движется беспорядочно. Так, личинки стрекоз предпочитают животных, совершающих зигзагообразные движения, а солнечная рыба скорее нападет на искусственную рыбку, если та извивается, чем на плавно движущийся объект. Иногда имеет значение соотношение формы и движения. Так, когда жабам предъявляют темную движущуюся полосу на белом фоне, они сразу же нападают на нее, когда полоса перемещается вдоль своей оси подобно червяку.
Узнавание добычи по форме. В ряде поведенческих работ Робинсон давал в неволе насекомых тамаринам. Выяснилось, что для опознания добычи тамаринам важны головы и конечности насекомых. Чтобы спастись от хищника, насекомые-жертвы выработали разнообразные защитные приспособления, скрывающие конечности и искажающие очертания тела.
Робинсон обнаружил, что двусторонняя симметрия, вероятно, служит одним из признаков добычи, на который обычно ориентированы хищники.
Те же принципы применимы и к узнаванию жертвами хищников. Так, например, силуэт ястреба, движимый над утятами или гусятами, вызывает реакцию страха, когда движется только в определенном направлении. Это объясняется тем, что короткая шея и длинный хвост характерны для ястреба, а длинная шея и короткий хвост - признаки летящего гуся. Жабы избегают фигур, похожих на змею с поднятой головой. Пиявка, движущаяся толчками, воспринимается как добыча, если ее передняя присоска находится на почве, но если эта присоска поднята в воздух, жаба принимает пиявку за врага.
Хотя для изучения сенсорных процессов могут быть применены разные чисто поведенческие методы, физиологическое исследование тоже может давать ценные сведения о деятельности органов чувств и о типе информации, которую они посылают в мозг.
Но для того чтобы установить, как мозг использует такую информацию, требуется сочетание поведенческого и физиологического подходов. Такой подход был применен Эвертом и его коллегами в обширных исследованиях по опознанию добычи и врагов жабами.
Физиологические работы показывают, что некоторое опознание добычи происходит на уровне сетчатки. Леттвин и др. отводили электрическую активность от зрительного нерва лягушки, когда объекты двигались в поле зрения. Они обнаружили четыре типа ответов, которые, по-видимому, соответствуют четырем типам ганглиозных клеток в сетчатке,являющихся детекторами:
1) неподвижной границы,
2) темного выпуклого движущегося объекта,
3) изменения контраста или движения
4) затемнения.
Таким, образом, информация, поступающая в мозг, включает угловой размер и скорость движения объекта, степень контраста с фоном и общий уровень освещенности. Однако, чтобы распознать добычу, жабе этой информации недостаточно.
Сетчатка одного глаза проецируется топографически на поверхностные слои противоположной зрительной покрышки. Электрическая стимуляция покрышки у свободно движущейся жабы приводит к ориентировочной реакции по направлению к соответствующей части поля зрения, т.е. вызывает такое поведение, как будто соответствующая часть зрительной покрышки стимулируется видом добычи. Электрическая стимуляция проекции сетчатки на таламической претектальной области вызывает у жабы реакцию избегания. Хирургическое разрушение части головного мозга приводит к тому, что животные начинают хватать любой движущийся предмет. Разрушение зрительной покрышки уничтожает всякую реакцию на движущиеся стимулы, в том числе и поведение избегания. Эти данные побудили Эверта постулировать, что проекция сетчатки на претектальную область таламуса вызывает поведение избегания, но что для этого также требуется некоторый приток возбуждения от зрительной покрышки. Проекция сетчатки на зрительную покрышку служит основой реакции схватывания всех движущихся объектов, но реакция на крупные или похожие на врага объекты тормозится претектальной областью таламуса и схватываются только мелкие объекты.
Гипотеза Эверта подтверждается физиологическими работами по изучению электрической активности нейронов в зрительной покрышке и претектальной области таламуса в ответ на стимуляцию сетчатки и других связанных с этой функцией частей головного мозга. Это исследование представляет большой интерес не только как демонстрация соответствующего участия сетчатки и мозга в фильтрации стимула, но так же как прекрасный пример того, чего можно достичь разумным сочетанием поведенческих и физиологических методов.
Зрительные приспособления к неблагоприятных условиям среды.
Многие различия между животными объясняются не столько их эволюционной историей, сколько экологическими условиями.
Животное, глаза которого приспособлены к яркому свету, будет обладать хорошей остротой зрения, восприятием цвета и движения. Такие типично дневные глаза относительно нечувствительны к низким уровням освещенности. Животные, адаптированные к сумеречному освещению, обладают более высокой чувствительностью, но в ущерб цветовому и детальному предметному зрению.
Видеть при слабом освещении нужно животным, ведущим ночной образ жизни, обитающим глубоко в воде и в пещерах. Эти условия не совсем сравнимы, потому что спектр приходящего света смещен в воде. Нет данных о том, что ночные животные обладают повышенной чувствительностью к длинным волнам. Однако некоторые из них владеют приспособлениями, повышающими чувствительность к свету. Глаза с широким зрачком и большим хрусталиком улавливают больше света, чем маленькие глаза. Глазами первого типа обладают, например, опоссум, домовая мышь и рысь. У других ночных животных, например сов и галаго, череп сужен с боков, что привело к цилиндрическому удлинению глаза. Цилиндрические глаза встречаются также у некоторых глубоководных рыб. Многие глубоководные животные специально адаптированы к господствующим условиям освещения. Максимумы поглощения у зрительных пигментов глубоководных рыб совпадают с длиной волны, максимально пропускаемой водой. У глубоко ныряющего кита северного плавуна зрительные пигменты сильнее всего поглощают более короткие волны, чем у неглубоко ныряющею серого кита. Точно так же глаза глубоководных морских ракообразных содержат зрительные пигменты с максимумами поглощения в более коротковолновой области, чем у мелководных ракообразных.
В дополнение к свойствам зрительных пигментов сумеречное зрение усиливают также другие специальные приспособления. Как правило, у ночных животных палочек больше, чем колбочек, а у некоторых из них, например у кошачьей акулы и галаго, мало или совсем нет. У многих ночных позвоночных близ фоторецепторов расположены отражающие свет образования, благодаря которым глаза "светятся", когда на них падает луч света. Свет, который проходит через фоторецептор не поглотившись, отражается обратно, что повышает вероятность поглощения.
Существует грань, за которой условия для зрения так тяжелы, что от глаз как от главного органа чувств приходится отказаться. У большинства животных, обитающих в пещерах, в глубине моря или же в мутной воде, глаза рудиментарные вследствие их регрессии. Например, пещерные земноводные обладают глазами на личиночной стадии, но лишены их во взрослом состоянии. Если этих животных воспитывать на свету, то у взрослых особей развиваются нормальные глаза. У молоди слепой пещерной рыбы имеются глаза, но у взрослых животных они дегенерируют. Садоглу обнаружил, что дегенеративное состояние глаз у пещерного вида определяется генами.
Среди млекопитающих кроты и летучие мыши являются самыми известными примерами дегенерации зрения. Очень маленькие глазки кротов у некоторых видов покрыты кожей. У плотоядных рукокрылых зрение развито хорошо, но у ночных летучих мышей, особенно у тех видов, которые ловят насекомых на лету, зрение очень слабое.
Органы чувств, заменяющие зрение.
Животные с очень слабым зрением, живущие при сумеречном свете, должны рассчитывать на другие органы чувств. Так, у придонных кошачьих сомов и ошибневых имеются разные сенсорные усики, которыми они ощупывают субстрат. Эти усики снабжены многочисленными осязательными рецепторами и хеморецепторами. Однако такие органы чувств не могут быть источником информации о величине и положении предметов в окружающей среде. Рыбы с нейромастами и органами боковой линии, чувствительными к вибрации, могут обнаруживать движущиеся предметы и получать некоторую информацию о неподвижных предметах по отраженным от них движениям воды. Но наилучшей заменой зрения животным служат электромагнитная чувствительность и особые слуховые приспособления.
Некоторые виды рыб используют электрическую чувствительность при обычной ориентации и коммуникации. Чувствительность к магнитным полям тоже обнаружена у ряда животных. Так, некоторые бактерии ориентируются к северному магнитному полюсу и реагируют на магнит в лабораторных условиях. С помощью электронного микроскопа у таких бактерий были обнаружены цепочечные структуры, содержащие кристаллы магнетита, которые также найдены в брюшке медоносных пчел и в сетчатке голубей. В Северном полушарии бактерии следуют наклонению магнитного поля Земли, и оно направляет их вниз, в анаэробный ил, их естественную среду обитания. В Южном полушарии у бактерий обратная полярность.
Рыбы используют электричество тремя разными способами.
1) Так называемые "сильноэлектрические" рыбы, такие, как электрический скат и электрический угорь, производят электрические разряды, способные оглушить жертву, но лишены электрической чувствительности.
2) Электрочувствительные рыбы, например Scyliorhinus и некоторые другие акулы, не производят электричества. Однако кошачьи акулы способны обнаружить даже зарывшуюся в песок добычу по локальному искажению электрического поля Земли. Для этого служат особые органы чувств - так называемые ампулы Лоренцини, широко распространенные по всей поверхности тела, особенно вблизи головы.
3) Так называемые "слабоэлектрические" рыбы генерируют свои электрические поля и чувствительны к электрическим изменениям среды. Обычно это ночные рыбы, обитающие в мутной воде. У них два типа электрочувствительных рецепторов: ампульные, которые отвечают на медленно изменяющиеся электрические поля, и клубневые, реагирующие только на быстрые их изменения. У некоторых видов один тип рецепторов, у других - оба. Эти рыбы генерируют слабые электрические поля посредством электрических органов, которые представляют собой видоизмененные мышцы или аксоны. Электрические разряды, как правило, испускаются с частотой до 300 импульсов в секунду. Некоторые рыбы способны менять частоту импульса. Когда одна рыба испытывает электрическое воздействие другой, она может изменить частоту своих импульсов, чтобы снизить это вмешательство. Электрорецепторы служат также для локализации предметов в окружающей воде по искажениям, которые эти предметы вызывают в электрическом поле. Некоторые рыбы, например Gymnarchus, отличают хорошие проводники от плохих, например металлический стержень от пластикового.
Слух заменяет зрение многим видам, и у некоторых из них образовались весьма интересные и специализированные добавления к нормальному слуху. Все эти адаптации способствуют точной локализации источника звука. Сравнение сигналов, приходящих в оба уха, служит главным способом локализации этого источника у позвоночных. Человек с одним ухом может, поворачивая голову, искать направление максимальной силы сигнала, потому что голова создает определенную звуковую тень. Два уха делают возможным одновременное сравнение, что позволяет локализовать источник гораздо быстрее и точнее. Если уши достаточно далеко отстоят друг от друга, то создается разница во времени прихода, в фазе звуковых волн, имеющих определенное направление, и их интенсивности. Таким образом, мелкие животные сравнивают только интенсивность звуков, а люди пользуются и монауральными, и бинауральными способами, и долгое время считалось, что они являются видом с наилучшей способностью к обнаружению источника звука. Но благодаря работе Пэйна, по слуховым способностям сова сипуха далеко превосходит человека. Она может локализовать и поймать свободно движущуюся мышь в полной темноте. Она даже способна определить направление движения животного и благодаря этому располагает свои когти вдоль длинной оси его тела. Сипуха пользуется низкочастотными компонентами звуков для локализации их источника в горизонтальной плоскости, а высокочастотными - для определения его положения по вертикали. Она не смешивает эти два типа информации, даже несмотря на то, что оба они основаны на сравнении звуков, приходящих в оба уха.
Самой совершенной заменой зрения является эхолокация, при которой животное испускает высокочастотные сигналы и обнаруживает предметы по возникающему от них эху.
Простые формы эхолокации встречаются у землероек, жирных козодоев и гималайских саланганов, которые ночуют и гнездятся в пещерах. Более совершенными ее формами обладают дельфины и другие морские млекопитающие, но своей вершины она достигает у летучих мышей.
В процессе эхолокации летучие мыши испускают залпы ультразвуковых импульсов малой длительности и высокой частоты. Такие короткие импульсы позволяют точно определять время образования эха, а значит, и расстояние до образующего его предмета. Звуки, производимые другими животными и ветром, обычно бывают низкочастотными, поэтому маловероятно, чтобы на ультразвуковые сигналы летучих мышей накладывались помехи. Однако, искусственные звуки частотой выше 20 кГц дезориентируют полет летучих мышей. Другое преимущество высоких частот состоит в возможности точной фокусировки, что делает возможным распознавание мелких объектов.
У летучих мышей много также специальных приспособлений, позволяющих им определять время и локализовать место возникновения эха от их ультразвуковых сигналов. У большинства рукокрылых, хватающих насекомых на лету, большие наружные уши, форма которых повышает дирекциональную чувствительность.
Зрительное опознание хищника и жертвы.
Большинство хищников встречается с множеством разных видов, которые служат им добычей и которых необходимо отличать от других организмов. При этом чаще всего используются три признака - величина, движение и форма.
Когда хищники должны выбирать между особями-жертвами, отличающимися только размерами тела, они обычно хватают самую крупную. Эта стратегия наиболее эффективна в отношении затрат энергии. Однако есть предел, за которым стимул больше не воспринимается как добыча. Например, когда обыкновенной жабе предъявляют как "добычу" объекты разной величины, она реагирует положительно на экземпляры, размеры которых находятся в определенных пределах, но активно избегает более крупные стимулы. Чтобы выбрать добычу определенной величины, жабе нужно оценить абсолютную величину видимого объекта, учитывая и его размеры на сетчатке, и расстояние до него. Жабы-повитухи в процессе развития постепенно научаются выбирать добычу постоянного размера.
В некоторых случаях наблюдается склонность к более крупной добыче, но не из-за активного предпочтения, а потому, что она лучше различима. Так, радужные форели чаще ловят крупных ракообразных, чем мелких, потому что крупные видны на большем расстоянии.
Для того чтобы узнать добычу, некоторым видам, например лягушкам и жабам, нужно, чтобы она двигалась. Обыкновенная каракатица в норме нападает только на движущихся креветок. Но если у нее отнять только что пойманную и парализованную ею креветку, то это головоногое немедленно снова нападет на нее даже на неподвижную. Некоторые хищники предпочитают добычу, которая движется беспорядочно. Так, личинки стрекоз предпочитают животных, совершающих зигзагообразные движения, а солнечная рыба скорее нападет на искусственную рыбку, если та извивается, чем на плавно движущийся объект. Иногда имеет значение соотношение формы и движения. Так, когда жабам предъявляют темную движущуюся полосу на белом фоне, они сразу же нападают на нее, когда полоса перемещается вдоль своей оси подобно червяку.
Узнавание добычи по форме. В ряде поведенческих работ Робинсон давал в неволе насекомых тамаринам. Выяснилось, что для опознания добычи тамаринам важны головы и конечности насекомых. Чтобы спастись от хищника, насекомые-жертвы выработали разнообразные защитные приспособления, скрывающие конечности и искажающие очертания тела.
Робинсон обнаружил, что двусторонняя симметрия, вероятно, служит одним из признаков добычи, на который обычно ориентированы хищники.
Те же принципы применимы и к узнаванию жертвами хищников. Так, например, силуэт ястреба, движимый над утятами или гусятами, вызывает реакцию страха, когда движется только в определенном направлении. Это объясняется тем, что короткая шея и длинный хвост характерны для ястреба, а длинная шея и короткий хвост - признаки летящего гуся. Жабы избегают фигур, похожих на змею с поднятой головой. Пиявка, движущаяся толчками, воспринимается как добыча, если ее передняя присоска находится на почве, но если эта присоска поднята в воздух, жаба принимает пиявку за врага.
Хотя для изучения сенсорных процессов могут быть применены разные чисто поведенческие методы, физиологическое исследование тоже может давать ценные сведения о деятельности органов чувств и о типе информации, которую они посылают в мозг.
Но для того чтобы установить, как мозг использует такую информацию, требуется сочетание поведенческого и физиологического подходов. Такой подход был применен Эвертом и его коллегами в обширных исследованиях по опознанию добычи и врагов жабами.
Физиологические работы показывают, что некоторое опознание добычи происходит на уровне сетчатки. Леттвин и др. отводили электрическую активность от зрительного нерва лягушки, когда объекты двигались в поле зрения. Они обнаружили четыре типа ответов, которые, по-видимому, соответствуют четырем типам ганглиозных клеток в сетчатке,являющихся детекторами:
1) неподвижной границы,
2) темного выпуклого движущегося объекта,
3) изменения контраста или движения
4) затемнения.
Таким, образом, информация, поступающая в мозг, включает угловой размер и скорость движения объекта, степень контраста с фоном и общий уровень освещенности. Однако, чтобы распознать добычу, жабе этой информации недостаточно.
Сетчатка одного глаза проецируется топографически на поверхностные слои противоположной зрительной покрышки. Электрическая стимуляция покрышки у свободно движущейся жабы приводит к ориентировочной реакции по направлению к соответствующей части поля зрения, т.е. вызывает такое поведение, как будто соответствующая часть зрительной покрышки стимулируется видом добычи. Электрическая стимуляция проекции сетчатки на таламической претектальной области вызывает у жабы реакцию избегания. Хирургическое разрушение части головного мозга приводит к тому, что животные начинают хватать любой движущийся предмет. Разрушение зрительной покрышки уничтожает всякую реакцию на движущиеся стимулы, в том числе и поведение избегания. Эти данные побудили Эверта постулировать, что проекция сетчатки на претектальную область таламуса вызывает поведение избегания, но что для этого также требуется некоторый приток возбуждения от зрительной покрышки. Проекция сетчатки на зрительную покрышку служит основой реакции схватывания всех движущихся объектов, но реакция на крупные или похожие на врага объекты тормозится претектальной областью таламуса и схватываются только мелкие объекты.
Гипотеза Эверта подтверждается физиологическими работами по изучению электрической активности нейронов в зрительной покрышке и претектальной области таламуса в ответ на стимуляцию сетчатки и других связанных с этой функцией частей головного мозга. Это исследование представляет большой интерес не только как демонстрация соответствующего участия сетчатки и мозга в фильтрации стимула, но так же как прекрасный пример того, чего можно достичь разумным сочетанием поведенческих и физиологических методов.
Понятие об этологии, ее задачи, методы. Современное состояние этологии.
Этология (от греч. ethos – обычаи, нрав, характер) – биологическая наука, изучающая поведение животных в естественных условиях; уделяет преимущественное влияние анализу генетически обусловленных (наследственных, инстинктивных) компонентов поведения, а также проблемам эволюционного поведения. Тесно связана с зоологией, эволюционным учением, физиологией, экологией, генетикой.
Этология изучает психику животных.
Одна из основных задач этологии - изучение развития психики животных в процессах филогенеза и онтогенеза.
А.Н.Леонтьев в одной из последних своих работ ("Образ мира", 1979) писал, что жизнь и животных, и человека осуществляется в предметном мире, и приспособление к нему происходит как приспособление к связям наполняющих этот мир вещей, к их движению, изменению во времени. Этология должна изучать характеристики образа мира, как специфичные только для животных, так и общие для животных и человека".
В этологии и раньше и теперь преобладает тенденция к изучению отдельных психических функций и способностей. Накоплены и обобщены этологические данные о различении свойств предметов животными, о выработке двигательных навыков, о способности к абстрагированию, об ориентации животных в пространстве и т.д. Тем не менее представление о психике конкретного вида животных или о психике той или иной особи остается очень мозаичным, похожим на лоскутное одеяло.
В настоящее время должен быть сделан переход от традиционного рассмотрения изолированных психических характеристик к анализу целостного поведения животного в природной среде обитания. Традиционно изучение психики животных происходит в условиях лабораторного эксперимента, которые далеки от природы. Часто задачи, которые ставят исследователи перед животными, искусственны, далеки от жизненных ситуаций; сама логика их постановки порой антропоморфна, то есть навеяна особенностями функционирования психики человека, а не психики животных. (Для животных иногда адаптивнее отказаться от решения задачи, чем пытаться ее решить).
Задачи и цели этологии
Задачи и цели проведения этологических наблюдений могут быть весьма разнообразны и направлены как на решение чисто практических, прикладных вопросов (определение физиологического состояния по внешним проявлениям или исследование возможности оставить новорожденного детеныша с матерью), так и решение теоретических проблем этологии (например, описание и последующее межвидовое сравнение поведенческих репертуаров или исследование механизмов парного взаимодействия).
При выборе конкретной задачи можно отталкиваться от какой-то проблемы или круга проблем и выбирать объект наблюдений так, чтобы наиболее легким путем разрешить поставленный вопрос. Этот подход считается наиболее продуктивным, однако на практике исследователь далеко не всегда обладает свободой выбора объекта. Поэтому чаще применяют противоположный подход - от объекта к проблеме, при этом усилия направляются на решение более легких вопросов. Если работа носит практический характер, то часто как объект, так и задачи исследования бывают жестко заданы. Основные направления этологических исследований, основной метод которых - наблюдения за поведением животных в неволе:
Выявление индивидуальных и видовых особенностей поведения
В рамках этого направления возможно сравнение структурных и временных характеристик различных поведенческих проявлений (выразительных движений, поз, поведенческих последовательностей, мимики, звуков), а также сравнительный анализ реакций на какие-либо стандартные ситуации. При изучении индивидуальных различий достаточно показать отличия выбранных показателей поведения сравниваемых животных; для выявления видовой специфики этого мало - необходимо также сравнивать пределы варьирования каждого признака в рамках одного вида, т.е. необходимы данные о большом числе животных.
Факторы, формирующие активность животных в неволе
Пользуясь тем, что в условиях неволи возможен сравнительно полный контроль действующих на животное внешних факторов, можно выделить то влияние, которое оказывает каждый из этих факторов. Часто такие наблюдения обладают большой практической ценностью. Обычно выделяют такие факторы, как: метеоусловия, режим кормления и уборок, режим и характер контактов с людьми, контакты с другими животными, нерегулярные воздействия (громкие шумы, появление необычных объектов и т.п.). В число внешних факторов входит и характер представленного животным пространства, хотя изучение пространственных характеристик активности обычно требует специальных наблюдений.
Описание тех сторон жизнедеятельности, которые малодоступны для изучения в природе
Здесь в первую очередь идет речь о поведении, связанном с размножением и выращиванием детенышей. Основные вопросы: поведенческие показатели готовности к размножению, описание процесса формирования пары, ухаживания, спаривания. Особое внимание необходимо уделить временным параметрам, а также регистрации аномальных поведенческих проявлений. При описании выращивания молодняка надо отмечать частоту и длительность кормлений, длительность пребывания самки в контакте с детенышами (и изменения обоих этих показателей по мере взросления молодняка), изменения в поведении детенышей.
Описание взаимоотношений с особями своего вида
Изучение механизмов внутривидового (социального) взаимодействия - одна из наиболее сложных и увлекательных задач этологии. При изучении этого вопроса в неволе появляется возможность помимо традиционных характеристик поведения (выразительных движений и поз, общего характера взаимодействий) анализировать такие характеристики социального взаимодействия, как динамика индивидуальных дистанций, степень синхронизации активности.
Методы этологии
Этологическому эксперименту должно предшествовать наблюдение за животными в природных условиях. Предпочтительнее эксперимент в природных условиях. Оптимальный вариант - наблюдение за повседневной жизнедеятельностью животных в условиях естественного эксперимента, поставленного самой природой. Допустимы исследования в искусственно созданных экспериментатором ситуациях, при условии их экологической адекватности
Этологические методы предполагают целостный, аутэкологический и, для таксонов позвоночных животных и некоторых беспозвоночных, - синэкологический подход. Аутэкология изучает связь отдельной особи со средой обитания. Синэкологический подход шире аутэкологического. Синэкологический подход предполагает, что особь живет не "сама по себе", а в популяции и, более того, в коадаптивном комплексе близких видов. Коадаптация - совместная адаптация (приспособление).
Различные подходы и основания для классификации поведения животных.
Существующие в настоящее время системы классификации поведения многообразны, так как число критериев, которые могут быть положены в ее основу, практически безгранично.
Классификация Д. Дьюсбери (1981), частично переработанная авторами, подразделяет поведение на три основные группы — индивидуальное, репродуктивное и социальное.
Индивидуальное поведение включает разнообразные акты, направленные на выживание и жизнеобеспечение отдельной особи:
- Локомоция - перемещения животного в пространстве, необходимые для выполнения практически любых приспособительных функций.
- Манипуляционная активность - совокупность действий особи с предметом, направленная на его адекватное использование в приспособительной деятельности. Составляет необходимый компонент пищевого, гнездостроительного, исследовательского, орудийного, а также других сфер поведения животного. Представляет собой систему определенным образом интегрированных элементов разного иерархического уровня. Сложность этой системы определяется не только морфологическими особенностями эффекторов (например, развитие кисти у обезьян или превращение передних конечностей в крылья у птиц), но также и общим уровнем организации поведения и психики животного (Дерягина, 1986).
- Исследовательская активность - комплекс реакций, которые знакомят животное с окружающей средой или источником раздражения и создают основу для «индивидуального программирования поведения» (Тинберген, 1963).
- Кормовое (или пищедобывательное) поведение - сложный, иерархически организованный многоуровневый комплекс двигательных актов, направленных на отыскание, схватывание, удержание добычи и последующее манипулирование с ней. В кормовом поведении действия с наследственно обусловленной видоспецифической программой тесно переплетены с действиями, приобретенными в результате индивидуального приспособления к среде. На пищедобывательных реакциях основано большинство методик изучения высшей нервной деятельности животных.
- Поведение, направленное на поиск оптимального температурного режима, которое обеспечивает процессы терморегуляции.
- Защитное поведение, связанное с поисками убежища, избеганием опасностей, охраной детенышей.
- Гигиеническое поведение направлено на поддержание чистоты тела, а также осуществление уринации и дефекации.
- Игра - совокупность специфически ювенильных проявлений всех форм поведения взрослого животного, характерная главным образом для молодых особей (Фабри, 1976; 1993), или та форма деятельности, «в которой складывается и совершенствуется управление поведением на основе ориентировочной деятельности» (Эльконин, 1997).
- Орудийная деятельность - особая категория индивидуального поведения, когда одни предметы окружающей среды используются для воздействия на другие в качестве средств, повышающих эффективность поведения в какой-либо сфере жизнедеятельности или даже уровень всего поведения в целом (Фабри, 1980). Это, несомненно, важная категория поведения, особенно в связи с проблемой разума животных. Однако она не столь универсальна, как рассмотренные выше, потому что к использованию орудий прибегают относительно немногие животные, причем в определенных и достаточно редких ситуациях.
Репродуктивное поведение связано с образованием брачных пар, выведением потомства и его воспитанием (его рассмотрение не входит в нашу задачу).
Социальное поведение включает все типы взаимодействий животных в сообществе, диапазон которых очень широк. Одни животные ведут исключительно одиночный образ жизни, большинство других образует разнообразные по численности (до многих тысяч особей) и уровню сложности сообщества. Многие виды в течение года переходят от одиночного к общественному образу жизни. Такие переходы характерны для многих видов птиц, некоторых грызунов и хищных млекопитающих (волки).
Проявление всех форм поведения находится под влиянием суточных, сезонных и других биологических ритмов.
Другие классификации поведения. Наиболее часто употребляемые классификации поведения подробно рассмотрены в фундаментальном руководстве Р. Хайнда «Поведение животных» (1975, гл. 2). Назовем некоторые из них.
- По непосредственным причинам, вызывающим тот или иной поведенческий акт. Например, все виды активности, частота или интенсивность которых достоверно повышаются под действием мужского полового гормона, можно объединить и обозначить как «половое поведение самца». Подобно этому все виды взаимодействия самцов с соперниками могут быть описаны как «агонистическое поведение».
• По функциям - группировка различных форм поведения в зависимости от той роли, которую они играют в жизни животного. Этот способ позволяет выделить такие категории, как угроза, ухаживание, добывание пищи и т. д.
• По происхождению в филогенезе - широко используется этологами при рассмотрении проблем эволюции поведения, в частности эволюции собственно инстинктивных действий. В ее основе лежит тщательное сравнительное изучение поведения видов с разной степенью родства.
• По способу формирования в онтогенезе - одна из важнейших и наиболее распространенных. Учитывает особенности проявления данного поведенческого акта в процессе индивидуального развития. Согласно этой классификации поведение традиционно подразделяют на врожденное (инстинктивное) и приобретенное в результате научения. Это приближается к принятому в физиологии ВИД выделению двух видов деятельности - условно- и безусловно-рефлекторной. Врожденное поведение обеспечивает приспособление особи к условиям среды, обычно характерным для вида в целом. Приобретенное поведение - способ индивидуального приспособления особи к меняющимся условиям среды.
Классификация форм поведения, предложенная Л. В. Крушинским.
Для точного ответа на вопрос о соотношении врожденных и приобретенных компонентов в поведении требуется специальный анализ с применением генетических методов и депривационных экспериментов (воспитание в изоляции от действия тех или иных факторов внешней среды). Ответ в каждом конкретном случае особый, причем наибольшие трудности возникают, когда речь идет о сложных когнитивных функциях (соотношение влияний генотипа и среды на психические особенности человека, а также на некоторые признаки поведения животных.
Зачастую само деление на «врожденное» и «приобретенное» производится совершенно неправомерно. Например, во многих случаях, когда поведенческий акт сформировался без явного участия ассоциативного обучения, его относят к категории врожденных. Однако это далеко не всегда верно, поскольку, во-первых, не все индивидуальные приспособительные поведенческие реакции есть результат обучения, и, во-вторых, если для появления поведенческого акта не требуется обучения, это еще не значит, что он осуществляется по готовой генетической программе. Здесь мы сталкиваемся с довольно распространенным вариантом смешения понятий.
Классификация форм поведения, предложенная Л. В. Крушинским (1986), соединяет в себе два критерия: 1) способ формирования в онтогенезе, 2) принципиальные нейробиологические механизмы, лежащие в основе осуществления данного поведенческого акта. Используя эти критерии, Л. В. Крушинский выделил три основные категории поведенческих актов.
* Поведение, которое строится по наследственно обусловленной программе и не требует для своего развития специального обучения или тренировки. В целом соответствует врожденным, или инстинктивным, действиям.
* Поведение, которое формируется постепенно, по мере накопления индивидуального опыта. Это различные формы привыкания и обучения.
* Поведение в новой для животного ситуации, на основе экстренного принятия им решения, без предварительного обучения и при отсутствии соответствующей наследственной программы. К этой категории относится элементарная рассудочная деятельность (мышление) животных.
Реальное поведение животного представляет собой сложное переплетение названных компонентов. В ряде случаев сходные по внешнему выражению действия могут различаться по их соотношению.
Индивидуальное приспособление животного к условиям среды может осуществляться двояко: обучение дает ему возможность приспособиться к постоянно действующим, несколько варьирующим, но знакомым факторам среды, а благодаря различным видам мышления, или рассудочной деятельности, животное может ответить экстренной и адекватной реакцией на непредвиденные изменения привычных условий, что практически невозможно достичь лишь на основе навыков и привычек, приобретенных научением.
Как подчеркивает Л. В. Крушинский (1986), особая приспособительная роль элементарной рассудочной деятельности состоит в «обеспечении адаптивной реакции уже при первой встрече с новой ситуацией».
Инстинкт и инстинктивное поведение: соотношение понятий.
Оксфордский толковый словарь по психологии
Инстинкт (лат – instinctus) - означает побуждавшим вынуждающий, при этом подразумевается, что такие импульсы естественны или врожденны. Существует четыре общих, различающихся между собой значения этого термина:
Неприобретенная реакция, характерная для членов данного биологического вида.
Тенденция или предрасположенность реагировать определенным способом, который является характерным для определенного вида. Эта предрасположенность – предполагаемая основа наблюдаемого поведения.
Сложный, скоординированный набор действий, универсального или почти универсально, проявляющийся у данного вида, возникающих при определенных стимульных условиях, определенных состояниях наличия влечения и определенных условиях развития. Это значение встречается прежде всего в этологии; см., например, врожденный механизм высвобождения, фиксированный паттерн активности и связанные с ними термины.
Любая из ряда неприобретенных, наследственных тенденций, которые, как предполагается, выполняют функции мотивационных сил сложных человеческих моделей поведения. Это значение, конечно, выражается в классическом психоанализе.
В фактическом использовании, способ применения этого термина резко отличается от теории к теории. Первой школой психологии, которая сделала инстинкт центральным понятием, был Фрейдизм. В своих ранних трудах Фрейд выделил два класса инстинктов: эго-инстинкты, или инстинкты самосохранения, и половые, или репродуктивные, инстинкты, позднее он ограничил их до Танатоса (инстинктов смерти) и Эроса (инстинктов жизни). Фрейд ясно показывал, что инстинкты были существенными мотивационными силами поведения но не определяли специфические поведенческие проявления. Основной акцент делался скорее на том, что является инстинктивным, чем на самих инстинктах.
В гормической психологии Макдугалла все поведение рассматривалось как преднамеренное или целенаправленное и мотивированное основными специфически видовыми наклонностями, инстинктами. В отличие от Фрейда, Макдугалл применял этот термин широко по отношению ко всем мотивационным конструкциям, и полученное в результате быстрое увеличение числа инстинктов подорвало научный базис теории.
Употребление этого термина этологами (прежде всего в значении 3, но во взаимодействии с 1 и 2) подчеркивало специфически видовые и биологические аспекты. Это становится ясным из определения Лоренца: "Поведение, которое в значительной степени определяется нервными механизмами, включенными в филогенез вида". Обратите внимание, что эта модель употребления специально включает поведение, в противоположность психоанализу, сосредоточивавшему внимание на мотивации.
Путаница во многом возникла из нескольких неразрешенных проблем:
а) Проблема наследственности: степень, в которой инстинкты являются биологически запрограммированными или следуют из факторов, относящихся к окружающей среде.
б) Видовая специфичность: являются ли инстинкты общими мотиваторами или они могут развиваться только внутри отдельных видов? Накапливающиеся свидетельства здесь имеют тенденцию поддерживать этологический подход, в котором основное внимание уделяется внутривидовому анализу.
в) Поведенческая специфика: степень, в которой определенные модели поведения должны включаться в понятие. Чем специфичнее предполагаемая модель поведения, тем больше возможность, что этот термин будет принимать недопустимую неопределенность.
В современных научных исследованиях все чаще избегают употребления понятия «инстинкт» ввиду многообразия и нечеткости его толкования. Понимаемое как врожденное, наследственно фиксированное, видотипичное, «инстинктивное поведение» складывается из инстинктивных действий или актов, которые в свою очередь состоят из отдельных инстинктивных движений (или поз, звуков и т.п.). Четкое разграничение этих терминов необходимо для анализа обсуждаемой категории поведения животных. Инстинкт - это совокупность врожденных компонентов поведения и психики животных и человека, а инстинктивное поведение животных совокупность сформировавшихся в процессе развития данного вида животных наследственно закрепленных, врожденных, общих для всех представителей вида компонентов поведения, составляющих основу жизнедеятельности животных.
Когда говорится, что поведение является совокупностью функций внешних, «рабочих» органов животного организма, необходимо различать сами эти функции и их ориентацию во времени и пространстве. И то и другое происходит на инстинктивной основе. Научением же можно только менять ориентацию этих функций. Это означает, что никаким научением нельзя заставить функционировать органы животного иначе, чем это обусловлено их генетически фиксированным строением. Нельзя «научить зайца спички зажигать», так как у него нет соответствующих морфофункциональных особенностей в строении конечностей. Но можно путем дрессировки научить зайца пользоваться своими конечностями естественным (инстинктивным) образом в нужный момент и в определенном направлении, т.е. ориентировать его инстинктивные движения во времени и пространстве путем научения.
Можно сказать, что вся жизнедеятельность животного организма, проявляющаяся во внешней активности, зиждется на инстинктивных движениях и других инстинктивных реакциях (термических, электрических, изменения окраски, выделения секретов и т.п.). эти реакции, главным образом движения, обеспечивают все жизненные функции организма, процессы обмена веществ, а тем самым существование особи и размножение. Вот почему мы говорим о примате инстинктивных движений и по отношению к нервной деятельности, сенсорике, психическому отражению, которые служат у животных лишь для осуществления этих движений, для их ориентации. Поэтому в эволюционном плане развитие психики являлось необходимым следствием (а затем уже - и предпосылкой) повышения уровня обмена веществ и двигательной активности.
Инстинктивное поведение не исчерпывается, однако, самими функциями экзосоматических органов, а включает в себя и механизмы их регулировки и пространственно-временной ориентации. В этом отношении регулировка и ориентация на основе научения, служат хотя и важным, но все же лишь дополнением к этим инстинктивным процессам. Примером инстинктивного поведения может служить сохранение способности к спариванию у самцов макаки или у крыс. Содержавшихся после рождения в изоляции. Развивавшиеся изолированно от других обезьян самцы макаки могли спариваться, но испытывали существенные затруднения. С другой стороны, самцы крыс, выращенные изолированно, могли спариваться даже легче и чаще, чем особи, развивавшиеся среди взрослых крыс. Эти примеры показывают, что даже сложное половое поведение может передаваться по наследству. Следовательно, инстинктивными принято называть такие формы поведения, которые передаются из поколения в поколение при помощи генетических механизмов. Эти формы поведения проявляются спонтанно или при появлении раздражителей, которые их вызывают.
Не только половое поведение может носить инстинктивный характер. Стрекотание самцов сверчков при исполнении брачной песни носит строгий видовой характер и неповторимый звуковой рисунок. Если провести скрещивание сверчков с разными песнями, то их потомки в первом поколении будут обладать песней смешанного звучания.
Особенно много примеров, связанных с врожденными навигационными способностями птиц. Почти все птицы, мигрирующие на значительные расстояния, ориентируются по звездному небу. Эта способность инстинктивна и передается по наследству.
В поведении птенцов выводковых птиц встречаются уникальные примеры наследуемого поведения. Птенцы австралийской кустарниковой индейки обладают одной из наиболее сложных форм врожденного поведения. Они вылупляются из яиц в сорной куче на глубине 60 см. Птенцы индейки не выклевываются из яйца, а разламывают скорлупу, упираясь в нее ногами и спиной. Затем они отдыхают несколько часов и начинают выбираться наружу без помощи родителей. Для этого птенцы ложатся на спину и разрывают материал кучи над собой. Материал падает с «потолка» камеры, а птенцы утрамбовывают его спиной «на полу». Подъем на поверхность кучи продолжается в течение 2,5 суток. Птенцы появляются на свет полностью оперенные и готовые вести самостоятельную жизнь. Следовательно, птенцы обладают довольно сложным врожденным поведением, которое они реализуют до первой встречи с представителями своего вида или родителями.
Среди наследуемых форм поведения представляет большой интерес феномен «самооплевывания» у ежей. Эта форма поведения ежей состоит в том, что ежи оплевывают иголки боков и крестцовой области после обследования незнакомого объекта. «Самооплевывание» является врожденной формой поведения и проявляется у детенышей в первые дни жизни. Это генетически передаваемое поведение выполняет информационную функцию и сохраняется в течение всей жизни.
Для того чтобы инстинктивное поведение могло проявиться, нужна не только стимулирующая ситуация вовне, но и определенные внутренние факторы в форме потребностей или мотиваций.
Внутренняя среда животного организма непрерывно обновляется, но эта среда остается по своим физиологическим показателям постоянной. Стабильность внутренней среды - совершенно необходимое условие для жизнедеятельности организма. Любые, даже незначительные, отклонения от нормы воспринимаются системой интерорецепторов и приводят в действие физиологические механизмы саморегуляции, в результате чего эти нарушения устраняются. Такими механизмами саморегуляции отечественный физиолог академик П.К. Анохин считал сложные динамические структуры, функционирующие по принципу обратной связи и обозначенные им как функциональные системы. Итак, первопричиной и основой мотивации поведения являются более или менее значительные и длительные отклонения от нормального уровня физиологических функций, нарушения внутренних ритмов, обеспечивающих жизнедеятельность организма. Эти сдвиги выражаются в появлении потребностей, на удовлетворение которых и направлено поведение.
Первостепенное значение имеют для внутренней мотивации поведения ритмические процессы, происходящие в центральной нервной системе. Собственная ритмика ее стволовой части у позвоночных и брюшных нервных структур у беспозвоночных обеспечивает прежде всего ориентацию поведения во времени. Сейчас хорошо известны автономные, самовозбуждающиеся колебательные процессы («внутренние» или «биологические часы»), которые регулируют общую ритмику жизнедеятельности организма. В отношении поведения это означает, что периодические колебания внешней активности животных, начало и окончание ритмично повторяющихся действий определяются ритмом «внутренних часов», синхронизированным с космическим временем. Существенные поправки или изменения вносятся в поведенческие ритмы многообразными биологически важными факторами среды, но общая "канва" инстинктивного поведения определяется самовозбуждающимися колебательными процессами с периодом, равным приблизительно суткам (околосуточный, «циркадный» ритм).
Эксперимент Шлейдта (установил, что клокотанье индюка вполне закономерно повторяется периодически даже в том случае, когда птица полностью изолирована от внешнего мира и даже лишена слуха) показывает, что эндогенные процессы в нервной системе способны обусловливать выполнение определенных инстинктивных движений и при полном отсутствии адекватных внешних стимулов. Так, немецкий нейрофизиолог Э. Хольст обнаружил в стволовой части головного мозга курицы ряд зон, активация которых (электрическим током) вызывает типичные инстинктивные движения различного функционального значения. При этом оказалось, что при раздражении одного и того же участка мозга с усилением силы раздражения одно инстинктивное действие сменяется другим в естественной последовательности. Учитывая, что все это происходило в отсутствие каких-либо адекватных внешних раздражителей, становится ясным, что на сугубо эндогенной основе могут выполняться не только отдельные инстинктивные движения, но и целые системы таких движений - инстинктивные действия.
Среди внутренних факторов инстинктивного поведения большую роль играют гормоны. Гормоны влияют на поведение в большей степени, чем любая другая система организма, за исключением мозга. Наибольшему гормональному контролю подвержено половое созревание, половой деморфизм, размножение, ухаживание, копуляция и инверсия пола. Особую роль играют гормоны в становлении половых различий в поведении. Развитие рассматривается как следование в определенном направлении, приводящее к становлению поведения, типичного для одного или другого пола. Тем не менее, не следует недооценивать влияние на поведение животного сложных социальных взаимодействий.
Специальные исследования мозга показали, что он организован одинаково по сравнению с двуполыми видами и способен адекватно отвечать на различные гормональные влияния. Введение в мозг ящериц андрогенов или прогестерона стимулировало передний гипоталамус и преоптическую область, что вызывало «псевдосамцовое» поведение. Введение эстрогена активизировало вентромедиальное ядро гипоталамуса и вызывало поведение, характерное для самок. Из этого примера понятно, что гормональная регуляция является основой сложного полового поведения.
Таким образом, поведение в своей основе внутренне так же организовано, так же зиждется на системах биологической саморегуляции, также закодировано в генетическом фонде вида, как и процессы, определяющие другие функции организма. В этом и проявляется единство всех форм жизнедеятельности животных.
Лишь в случае объединения внешних и внутренних факторов может произойти запуск инстинктивного поведения. В результате создается впечатление, что животное специфически реагирует на какую-то определенную стимуляцию в определенный момент. Такое соответствие между типом раздражения и типом реакции навело этологов на мысль о существовании какого-то механизма, способного решать, какую именно из всех свойственных данному виду форм поведения следует запустить в том или ином случае. Этот механизм, присущий данному виду и встроенный в его мозг с самого рождения, получил название врожденного пускового механизма. Его можно было бы сравнить с заложенной в вычислительную машину программой, которая с учетом информации, поступающей извне, позволяет решить, в какой момент и в каких условиях нужна та или иная ответная реакция.
Инстинкт. Врожденный пусковой механизм.
Поведение в своей основе внутренне так же организовано, так же зиждется на системах биологической саморегуляции, также закодировано в генетическом фонде вида, как и процессы, определяющие другие функции организма.
Лишь в случае объединения внешних и внутренних факторов может произойти запуск инстинктивного поведения. В результате создается впечатление, что животное специфически реагирует на какую-то определенную стимуляцию в определенный момент. Такое соответствие между типом раздражения и типом реакции навело этологов на мысль о существовании какого-то механизма, способного решать, какую именно из всех свойственных данному виду форм поведения следует запустить в том или ином случае. Этот механизм, присущий данному виду и встроенный в его мозг с самого рождения, получил название врожденного пускового механизма. Его можно было бы сравнить с заложенной в вычислительную машину программой, которая с учетом информации, поступающей извне, позволяет решить, в какой момент и в каких условиях нужна та или иная ответная реакция.
Подобного рода механизм управляет в большинстве случаев поведением очень многих видов. Особенно это относится к брачному поведению. Так, например, у гусей и уток с наступлением времени брачных церемоний самец начинает проделывать ряд строго определенных движений (приподнимает туловище, раскрывает крылья, виляет хвостом и т. п.) в определенном порядке в ответ на приближение или удаление самки. Число и интенсивность этих движений возрастают по мере того, как все больше проявляется рецептивность самки, и достигают кульминации при сближении и спаривании.
Под врожденным пусковым механизмом следует понимать совокупность нейросенсорных систем, обеспечивающих адекватность поведенческих актов по отношению к «пусковой ситуации»: настройку анализаторов на восприятие специфических раздражителей и распознавание последних, интеграцию соответствующих раздражителей и растормаживание (или активацию) нервных центров, связанных с данным поведенческим актом.
Понятие об импринтинге, его значение для животных и человека.
Импринтинг (англ. imprint - запечатлевать, оставлять след) - специфическая форма научения у новорожденных высших позвоночных, при котором в их памяти автоматически фиксируются отличительные признаки поведения первых увиденных ими внешних объектов. Чаще всего такими объектами становятся родительские особи, выступающие одновременно носителями типичных признаков вида, братья и сестры, пищевые объекты, в том числе животные-жертвы и др. Иимпринтинг возможен в течение определенного, обычно весьма ограниченного («сенсибильного», «критического») периода. Процесс импринтинга совершается чрезвычайно быстро, без внешнего подкрепления, и результат его, как правило, необратим. Он обеспечивает животным охрану потомства (следование детей за родителями), узнавание родителей, членов сообщества, сородичей, будущих половых партнеров, признаков местности и др. В искусственных, экспериментальных условиях у некоторых видов животных и птиц импринтинг может быть вызван любым живым или неживым движущимся объектом.
Воспринимая сигналы от родителей, каждая особь запечатлевает в своем сознании образ животных своего собственного вида, что и определяет впоследствии различные проявления его поведения в сообществе. Таким образом, полученная информация закрепляется навсегда в мозгу животного именно в тот период, когда возможность ошибок сведена до минимума, то есть когда забота о нем со стороны родителей осуществляется наиболее полно.
Уже много лет стоит вопрос, связанный с первоначальным пониманием импринтинга, согласно которому животные посредством этой формы обучения вырабатывают для себя обобщенный образ собственного вида. На сегодняшний день уже изучено множество случаев, поясняющих, каким образом даже при наличии полиморфизма животные находят себе пару, руководствуясь навыками, полученными в раннем возрасте. Это касается птиц, млекопитающих, рыб, а возможно, и насекомых. Сигналы, благодаря которым животные распознают себе подобных, в естественных условиях обычно одинаковы у всех особей данной популяции; если некоторые особи наделены какими-либо другими характерными отличительными признаками, то половое влечение по отношению к ним не срабатывает. Но иногда - а это проявляется главным образом среди домашних животных, у которых сигналы видового распознавания могут значительно варьировать, - наблюдаются случаи межвидового полового влечения.
Как правило, посредством импринтинга закрепляются реакции на те сигналы, которые в дальнейшем будут определять поведение животного по отношению к тому или иному члену данного вида (а также к определенному типу пищи или местообитанию). Ответные реакции на такие сигналы обычно зависят от предварительных навыков и возникают у животных непроизвольно, без обучения.
Существуют также такие формы импринтинга, когда запечатлевается не только объект влечения, но и само его поведение. Наиболее яркий пример этому демонстрируют птицы, среди которых действует так называемое голосовое запечатление. Немецкий этолог Николаи, воспитавший выводок снегирей, заметил, что во время критического периода его птицы необратимо усвоили нехитрый мотив, который он постоянно насвистывал, и стали использовать эту мелодию в качестве сигнала межвидового распознавания. Другие снегири, которые росли бок о бок с канарейками, научились петь по-канареичьи. Таким образом, ему удалось вывести семейство снегирей, которые посредством запечатления передавали из поколения в поколение эту способность петь по канареечьи.
Различные птичьи «диалекты» возникают в различных географических районах именно благодаря голосовому запечатлению. Таким образом, небольшие, но весьма характерные голосовые различия фиксируются и передаются затем другим поколениям. Если же такие различия сказываются к тому же в изменениях полового влечения, то тем самым они могут способствовать выделению из первоначального вида отдельных групп особей, все более обосабливающихся за счет их половой изоляции от других популяций данного вида. В результате могут возникнуть новые виды птиц.
Еще один случай запечатления, когда приобретение навыков, ограниченное по времени критическим периодом, не только порождает способность распознавать сигналы и инстинктивно отвечать на них, но и определяет многие другие повадки животных, а также их способность к звукоподражанию. Речь идет о детях, выращенных волками, либо воспитанных в условиях изоляции. Попытки перевоспитать такого ребенка, с младенческого возраста находившегося в полном отрыве от людей, почти всегда заканчивались неудачей, особенно если дело касалось подростков, которым удалось утвердиться и добиться определенного положения в вырастившей их стае животных. Руководствуясь этими фактами, Жан-Клод Арман, обнаружив мальчика-газель, предпочел наблюдать за мальчиком в той естественной обстановке, к которой он прекрасно приспособился.
Человек более чем любое другое живое существо способен приспосабливаться и усваивать условия той общественной среды, в которой он живет. Однако это отнюдь не означает отсутствия генетической предопределенности. Коль скоро человек способен создавать материальную культуру и вести себя в соответствии с полученным им воспитанием и на основе навыков, усвоенных при взаимодействии с окружающей социальной средой, то это объясняется тем, что такая способность унаследована генетически.
Было бы слишком смелым говорить о наличии импринтинга как такового у человека, но некая близкая аналогия с этим явлением все же существует. На примере мальчика-газели можно видеть, что он действительно усвоил повадки газелей: мальчик вел дневной образ жизни, а по ночам спал, свернувшись калачиком и тесно прижавшись к животным, поддерживал с ними общение голосом, а также потираясь головой об их морды или касаясь их языком. И все же нечто присущее только человеку было в нем. Мальчик щипал траву, но также ловко взбирался на вершину финиковых пальм за плодами. Он ходил на четвереньках, но, когда ему приходилось следовать за быстро удаляющимся стадом, он бежал на двух ногах.
Итак, в ходе импринтинга неизгладимо запечатлеваются некоторые повадки и половое влечение к живым существам, служившим объектами запечатления во время критического периода.
Если взаимно введенные в заблуждение виды сходны генетически, то может произойти гибридизация. Так, американскому орнитологу начала века Уитмену удалось вывести несколько гибридов различных пород голубей. Он подменивал яйца у голубей, подлежащих скрещиванию, и добивался того, что новое потомство приобретало некоторые характерные признаки поведения, свойственные смежной породе. Когда выведенные таким путем голуби подрастали, они начинали испытывать взаимное половое влечение и легко спаривались, давая гибридное потомство.
Естественные функции импринтинга могут быть весьма различны. Первая и самая прямая - это формирование привязанности между потомством и родителями. На основе этого развиваются различные особенности поведения в сообществе и полового влечения. Но существует также и другая форма запечатления, не связанная с жизнью в сообществе, когда некоторые виды животных обретают необратимые сведения о том, чем им надлежит питаться и где селиться. Лососи, например, способны отыскать дорогу из открытого моря к месту нереста, потому что еще мальками они научились распознавать химический состав родных водоемов и могут безошибочно выбрать обратный путь из своего первого миграционного путешествия.
Итак, импринтинг - это форма обретения устойчивых навыков, которые остаются неизменно запечатленными на протяжении всей жизни. Следовательно, главная функция импринтинга состоит в получении информации об окружающей среде и прочном закреплении полученных сведений. Эта форма обучения имеет явные отличия от других форм, которые предоставляют животным достаточную свободу приспособляемости к среде обитания в зависимости от переменчивости ее условий. Таким образом, импринтинг можно рассматривать как некую переходную форму от инстинкта к познанию.
Пищевой инстинкт. Пищевое поведение различных животных и современного человека. Регуляция пищевого поведения.
Пищевое поведение присуще всем представителям животного мира. В основе пищевого поведения лежит взаимодействие центральных механизмов возбуждения и торможения. Составные элементы этих процессов отвечают как за реакцию на различные пищевые раздражители, так и на характер движений при еде. Определенную роль в формировании пищевого поведения играет индивидуальный опыт животного, в частности опыт, определяющий ритмы поведения.
Начальная фаза пищевого поведения – это поисковое поведение, вызванное возбуждением. Поисковое поведение определяется лишением животного пищи и является результатом возрастания реактивности к внешним раздражителям. Конечная цель поискового поведения – нахождение пищи. В эту фазу животное особенно чувствительно к раздражителям, которые косвенным образом указывают на присутствие пищи. Признаки, служащие раздражителями, бывают общими для разных видов пищи или характеризуют конкретный ее вид, что чаще всего наблюдается у беспозвоночных. Например, для пчел таким раздражителем может служить окраска венчиков цветка, а для термитов – запах гниющей древесины. Все эти раздражители вызывают различные виды активности. В зависимости от обстоятельств и вида животного это может быть схватывание добычи, ее предварительная подготовка и поглощение. Например, у волков наблюдается определенный способ охоты на разные виды копытных, рысь же охотится на все виды добычи одинаково. У хищных млекопитающих существуют определенные «ритуалы» при поедании добычи. Ласка поедает мышевидных грызунов с головы, а когда добычи много, довольствуется только мозгом жертвы. Крупные хищники также предпочитают поедать жертву, начиная с мышц шеи и внутренностей.
Когда животное начинает насыщаться, обратные связи, вызванные раздражением рецепторов рта, глотки и желудка, сдвигают равновесие в сторону торможения. Этому способствует также изменение состава крови. Обычно процессы торможения опережают компенсаторные способности тканей и протекают с различной скоростью. У некоторых животных процессы торможения воздействуют только на завершающий акт пищевого поведения и не касаются поведения поискового. Поэтому многие сытые млекопитающие продолжают охотиться, что свойственно, например, куньим, некоторым крупным кошкам.
Существует множество разнообразных факторов, определяющих привлекательность различных видов пищи, а также количество поглощаемой пищи. Лучше всего эти факторы изучены на примере крыс. У этих грызунов, отличающихся сложным поведением, новизна пищи может служить фактором, способствующим как увеличению съеденной пищи, так и уменьшению ее количества. Обезьяны чаще едят новую для себя пищу в малых дозах, но если обезьяна замечает, что ее сородичи поедают эту пищу, количество съеденного заметно возрастает. У большинства млекопитающих первыми новую пищу пробуют молодые животные. У некоторых стайных млекопитающих и птиц отдельные особи чаще пробуют незнакомый корм, находясь в окружении сородичей, и очень осторожно относятся к нему, находясь в изоляции. Объем поглощаемой пищи может зависеть и от ее количества, имеющегося в наличии. Например, в осенний период медведи поедают груши в садах в заметно большем количестве, чем с отдельно стоящих деревьев.
К пищевому можно отнести такое широко распространенное поведение, как запасание корма. Для обеспечения кормом личинок насекомых оно сводится к деятельности по откладке яиц на живые объекты (оводы), проявлению паразитизма, деятельности жуков-могильщиков. Запасание корма широко распространено и среди млекопитающих. Например, корм запасают многие виды хищников, причем формы запасания у них чрезвычайно разнообразны. Домашняя собака может просто зарыть кусок мяса, оставшийся от обеда, а горностай, куница устраивают целые склады, состоящие из трупов мелких грызунов. Запасают корм и многие виды грызунов, у некоторых из них (хомяки, мешотчатые крысы) имеются специальные защечные мешки, в которых они переносят корм. У большинства грызунов сроки запасания пищи строго ограничены, в большинстве случаев они приурочены к осени, когда созревают семена, орехи, желуди.
Регуляция пищевого поведения осуществляется рядом структур ЦНС и прежде всего двух взаимодействующих центров – центром голода (латеральное ядро гипоталамуса) и центром насыщения (вентромедиальное ядро гипоталамуса). Электрическая стимуляция центра голода провоцирует акт еды у сытого животного, тогда как стимуляция центра насыщения прерывает прием пищи. Разрушение центра голода вызывает отказ от потребления пищи (афагия) и воды, что часто приводит к гибели животного. Электрическая стимуляция латерального ядра гипоталамуса увеличивает секрецию слюнных и желудочных желез, желчи, инсулина, усиливает моторную деятельность желудка и кишечника. Повреждение центра насыщения увеличивает прием пищи (гиперфагия). Практически сразу после такой операции животное начинает есть много и часто, что приводит к гипоталамическому ожирению. При ограничении пищи масса тела уменьшается, но как только ограничения снимают, вновь проявляется гиперфагия, снижающаяся лишь при развитии ожирения. Эти животные проявляли также повышенную разборчивость при выборе пищи, предпочитая наиболее вкусную. Ожирение, следующее за повреждением вентромедиального ядра гипоталамуса, сопровождается анаболическими изменениями: изменяется обмен глюкозы, повышается уровень холестерина и триглицеридов в крови, снижается уровень потребления кислорода и утилизации аминокислот. Электрическая стимуляция вентромедиального гипоталамуса уменьшает секрецию слюнных и желудочных желез, инсулина, моторику желудка и кишечника. Таким образом, можно заключить, что латеральный гипоталамус вовлечен в регуляцию метаболизма и внутренней секреции, а вентромедиальный гипоталамус оказывает на нее тормозное влияние.
В норме сахар крови является одним из важных (но не единственным) факторов пищевого поведения. Его концентрация весьма точно отражает энергетическую потребность организма, а величина разности его содержания в артериальной и венозной крови тесно связана с ощущением голода или сытости. В латеральном ядре гипоталамуса имеются глюкорецепторы (нейроны, в мембране которых есть рецепторы к глюкозе), которые тормозятся при увеличении уровня глюкозы крови. Установлено, что их активность в значительной степени определяется глюкорецепторами вентромедиального ядра, которые первично активируются глюкозой. Гипоталамические глюкорецепторы получают информацию о содержании глюкозы в других частях тела. Об этом сигнализируют периферические глюкорецепторы, находящиеся в печени, каротидном синусе, стенке желудочно‑кишечного тракта. Таким образом, глюкорецепторы гипоталамуса, интегрируя информацию, получаемую по нервным и гуморальным путям, участвуют в контроле приема пищи.
Получены многочисленные данные об участии различных мозговых структур в контроле приема пищи. Афагия (отказ от пищи) и адипсия (отказ от воды) наблюдаются после повреждения бледного шара, красного ядра, покрышки среднего мозга, черной субстанции, височной доли, миндалины. Гиперфагия (обжорство) развивается после повреждения лобных долей, таламуса, центрального серого вещества среднего мозга.
Несмотря на врожденный характер пищевых реакций, многочисленные данные показывают, что в регуляции приема пищи важная роль принадлежит условно-рефлекторным механизмам. Это является основной причиной переедания и, следовательно, приобретения лишней массы тела современным человеком. Вспомните, каким обжорством мы страдаем, приходя в гости. В регуляции пищевого поведения участвуют многие факторы. Общеизвестно влияние на аппетит вида, запаха и вкуса пищи. Степень наполнения желудка также влияет на аппетит. Хорошо известна зависимость приема пищи от температуры окружающей среды: низкая температура стимулирует прием пищи, высокая – тормозит. Конечный приспособительный эффект всех механизмов, участвующих в пищевом поведении, состоит в приеме количества пищи, сбалансированного по калорийности с расходуемой энергией. Этим достигается постоянство массы тела.
Регуляция полового поведения. Стратегии полов и социальная организация вида.
У млекопитающих гипофиз секретирует гонадотропные гормоны, которые оказывают регулирующее влияние на различные физиологические процессы, имеющие отношение к размножению. Наибольший эффект гонадотропные гормоны оказывают на функционирование половых желез. Гормоны мужских и женских половых желез (андрогены и эстрогены), действуя на ЦНС, активируют половое поведение. В организме взрослой женщины наблюдаются периодические изменения различных функций, связанные с повторяющимися процессами роста и созревания фолликулов, овуляции и образования желтых тел. Созревание фолликулов сопровождается высоким уровнем секреции половых гормонов. В этот период яйцеклетка освобождается из фолликула (овуляция) и попадает в половые пути. Этот период является самым благоприятным для зачатия. Важную роль здесь играет действие эстрогена на гипоталамус.
Удаление у крысы яичников вызывает снижение и прекращение половой активности. Введение эстрогена в гипоталамус самок с удаленными яичниками сопровождается всем комплексом проявлений эструса. Наибольшее значение для организации полового поведения самок, по‑видимому, имеет гипоталамус. Его разрушение приводит к потере половой активности. У кроликов стимуляция преоптической области и базальной части гипоталамуса вызывает овуляцию. Радиоактивно меченый эстроген в наибольшем количестве накапливается в переднем гипоталамусе и ядрах миндалины. При локальных повреждениях в различных областях гипоталамуса можно заблокировать либо поведение, характерное для эструса, либо секрецию гонадотропного гормона.
У самцов животных кастрация прекращает половую активность, хотя у отдельных видов, например у домашних копытных, половое влечение сохраняется в течение года. Восстановление дооперационного уровня половой активности достигается постоянным введением половых гормонов. Нередко введение самкам мужского полового гормона вызывает поведение, характерное для самцов.
Аналогичный эффект можно наблюдать у кастрированных самцов после введения эстрогена – поведение, характерное для самок.
В эмбриональном или раннем постнатальном периоде развития (в зависимости от вида животного) происходит половая дифференцировка головного мозга по отношению к регуляции секреции гонадотропных гормонов гипофиза, реактивности к половым гормонам, поведению, росту и др. Половые различия мозга определяются разными структурами и прежде всего медиальным гипоталамусом. Важным фактором указанной дифференцировки является действие на мозг в «критический период» развития организма половых гормонов. До этого мозг независимо от генетического пола эмбриона или новорожденного имеет женский тип. Под влиянием андрогена, вырабатываемого семенниками, мозг маскулинизируется, т.е. приобретает мужской тип. Если кастрировать новорожденных самцов крыс, мозг сохранит женский тип. Если таким животным пересадить яичник и вагинальную ткань, у них в дальнейшем устанавливается женский половой цикл. Описанный эффект не возникает при кастрации на 10‑й день, поскольку мозг уже маскулинизирован. Однократное введение новорожденной самке тестостеронпропионата (аналог тестостерона) приводит к маскулинизации мозга и, как следствие этого – к отсутствию овуляторного цикла (овуляции и развития желтых тел). В период половой дифференцировки мозга определяется также реактивность центров полового поведения к специфическим раздражителям и гормонам. У приматов, включая человека, половые отличия в функционировании мозга устанавливаются во внутриутробном периоде.
У млекопитающих спаривание представляет сложный процесс, в регуляции которого принимают участие разные отделы ЦНС. Большая часть вегетативных и двигательных реакций, составляющих половой акт, интегрируется на спинальном уровне в поясничных и крестцовых сегментах. После перерезки спинного мозга путем стимуляции половых органов можно вызвать эрекцию и даже эякуляцию. Данный эффект отсутствует у животных с перерезкой ствола мозга каудальнее варолиева моста. По‑видимому, это свидетельствует о торможении спинальных механизмов продолговатым мозгом.
Гипоталамус участвует в регуляции половой активности у самца. Стимуляция области медиального пучка переднего мозга и рядом расположенных участков гипоталамуса вызывает у обезьян комплекс эмоциональных и поведенческих проявлений готовности к спариванию. Миндалина также включена в систему контроля полового поведения. У обезьян и кошек после удаления миндалины и периформной коры развивается гиперсексуальность, проявляющаяся в попытках спариться с представителями своего пола, животными других видов и даже неодушевленными предметами. Возможно, в норме миндалина оказывает тормозное влияние на половое поведение. Значение коры больших полушарий в организации полового поведения в ходе эволюции млекопитающих возрастает. У приматов при становлении рефлекса спаривания обучение играет существенную роль. У других животных, например у кошек и собак, половое поведение после декортикации не утрачивается, но оно, как правило, не завершается спариванием.
Социальная организация у животных разных видов чрезвычайно различна: от простой кооперации между самцом и самкой до сложных сообществ, как у некоторых приматов. Одним из важнейших факторов, влияющих на социальную организацию вида, является система размножения данного вида. Для этого необходимо оценить альтернативные эволюционные стратегии, возможные для самцов и самок.
Стратегия полов
Основная эволюционная проблема для самцов привлечь самок при наличии конкуренции со стороны других самцов. Основная эволюционная проблема для самок - выбрать в качестве полового партнера такого самца, который обеспечит потомство наилучшими шансами на выживание и размножение. Самка с самого начала имеет преимущество, так как она может отказаться от спаривания. Самец немногого может добиться, если самка его не принимает. Казалось бы, самец может заставить самку спариваться, однако обычно это довольно трудно, так как у большинства видов самка должна принять особую позу, чтобы копуляция стала возможна. Однако у некоторых животных, в том числе у крякв и скорпионовой мухи (Panorpa), иногда наблюдается принудительная копуляция. Самец скорпионовой мухи во время ухаживания обычно приносит самке свадебный подарок, часто это мертвое насекомое, вытащенное из тенет паука. Копуляция происходит, пока самка ест этот подарок. Иногда самец принуждает самку к копуляции и без подарка. В этом случае при успешной принудительной копуляции самец оказывается в выигрыше, поскольку ему не нужно рисковать во время поисков свадебного подарка. Известно, что 65% взрослых самцов скорпионовой мухи гибнет в паутине. Однако степень успешности принудительного спаривания, очевидно, невелика, поэтому к такой стратегии насекомые прибегают в крайнем случае.
Самцы обычно остаются в выигрыше в каждом сексуальном взаимодействии с самкой, если спариваются с ней, тогда как самки не всегда выигрывают и могут даже поплатиться снижением приспособленности. Поэтому для самок наилучшая стратегия быть скромной. Джордж Уильяме описал ухаживание как спор между самцом, навязывающим товар, и самкой, не желающей его брать. Самке не надо было бы оставаться недоступной, если бы она прямо могла оценить приспособленность самца и его будущее поведение. Однако все, о чем самка может судить, это внешний вид самца и его поведение в настоящий момент. Самка должна заставить самца проявить свою истинную натуру или повернуть дело так, чтобы его интересы совпали с ее интересами. В обоих случаях наилучший способ поведения для самки - быть осторожной и продлить период ухаживания, чтобы удостовериться, что ее потенциальный партнер - самец того же вида. Естественный отбор благоприятствует особям с отчетливыми признаками и поведением, позволяющими идентифицировать их как представителей конкретных видов. Совокупность этих признаков и поведения играет очень важную роль в ухаживании и называется изолирующими механизмами, поскольку эти механизмы обеспечивают репродуктивную изоляцию между видами, снижая возможность гибридизации. Три примера изолирующих механизмов.
1. На арктическом побережье Канады обитают четыре близкородственных вида чаек. В некоторых местах все четыре вида сосуществуют в одной колонии, но при этом не скрещиваются друг с другом. Птицы чрезвычайно похожи и различаются только отметинами на концах крыльев и окраской окологлазничной области. Изменяя в эксперименте эти цветовые отметины, можно добиться спаривания разных видов.
2. Обширные исследования скрещивания различных пород домашних кур показали, что внешний вид птицы очень важен. Так во время ухаживания петух обходит курицу вальсирующими движениями и при этом демонстрирует оперение. При согласии спариваться куры припадают к земле в типичной позе готовности. После этого петух делает садку. Поскольку куры оценивают своих партнеров в основном по внешнему виду, а не по различиям в поведении, экспериментальное изменение различных типов окраски петухов уменьшало их привлекательность для кур.
3. Известно, что самцы гуппи при ухаживании не делают различия в выборе самки, тогда как самки довольно строго выбирают партнера, основываясь на окраске самца и его поведении при ухаживании. Проводя эксперименты в аквариуме, Лили обнаружил, что самцы видов Poecilia picta и Poecilia reticulata часто ухаживают и делают попытки спариться с самками и своего, и чужого вида. А готовые к спариванию самки принимали ухаживания самцов только своего, но не другого вида. Но, если у самок не было выбора, они спаривались и с самцами другого вида.
Самка должна сделать все, чтобы удостовериться, что она спаривается с половозрелым и полноценным в половом отношении самцом. Продлевая период ухаживания, она с большей вероятностью будет спариваться с самцом, испытывающим сильную мотивацию и мощным в половом отношении. У европейского обыкновенного тритона (Triturus vulgaris) интенсивность ухаживания самца зависит от числа сперматофоров (капсул со спермой), которые самец продуцирует во время данной встречи. Во время ухаживания самец откладывает сперматофор на дно водоема; вероятность того, что самка успешно подберет его, возрастает в процессе ухаживания. Чтобы отложить три сперматофора, самец должен оставаться под водой не дыша гораздо дольше обычного. Таким образом, самка оценивает приспособленность самца.
Оценка самца самкой облегчается, если самец может продемонстрировать, что он являет собой достойный выбор. Некоторые исследователи полагают, что в этом отношении важную роль играет ритуальное кормление. Ритуальное кормление, во время которого самец подносит самке пищу или напоминающий пищу предмет, наблюдается у многих видов. У серебристой чайки (Lams argentatus)самка выпрашивает пищу, как если бы она была птенцом, а самец отрыгивает пищевой комок, который самка затем съедает.
У некоторых видов ритуальное кормление формализовано и подношения реальной пищи не происходит. Ритуальное кормление влияет на вес кладки у обыкновенной крачки(Sterna hirundo) и может служить показателем будущего участия самца в выкармливании птенцов. Вирджиния Нибур (Niebuhr, 1981) установила, что ритуальное кормление у серебристой чайки было надежным показателем последующего участия самца в выкармливании птенцов. Оно также указывает на склонность самцов участвовать в насиживании и защищать птенцов после вылупления.
Наилучшей стратегией самки в этом случае будет просто продлить или усложнить процесс ухаживания, так как это затруднит спаривание самца с другими самками, в особенности если сезон размножения короткий или мало свободных самок; иными словами, самцы, затрачивающие много сил и времени в процессе деятельности, предшествующей спариванию, такой, как ухаживание, строительство гнезда, защита территории, будут располагать меньшими временными и энергетическими ресурсами, чтобы преследовать других самок. Менее вероятно, что самец, уже затратив так много усилий на одну самку, оставит ее, вынуждая, таким образом, заботиться о потомстве в одиночестве. Более того, если самец утратит возможность спариваться с другими самками, он сможет увеличить свою приспособленность, только вкладывая оставшиеся ресурсы в свое потомство. Доукинс (Dawkins, 1976) назвал это «ситуацией семейного счастья».
Нет никаких сомнений, что «семейное счастье» можно наблюдать у некоторых моногамных видов. Однако это не является эволюционно стабильной ситуацией при всех обстоятельствах, так как здесь возможно вмешательство самок, придерживающихся иной стратегии. Доукинс создал модель, в которой возможны две стратегии для самок - «скромная» и «легкомысленная» и две для самцов - «верный» и «волокита». Скромные самки требуют продолжительного ухаживания перед копуляцией; легкомысленные самки копулируют без предварительного ухаживания. Верные самцы добровольно ведут длительное ухаживание, а после спаривания помогают самке вырастить потомство. Самцы-волокиты долго не ухаживают и, если не удастся спариться вскоре после встречи с самкой, оставляют ее и ищут другую. Самцы-волокиты могут оставить самку и после копуляции.
В популяции, где доля легкомысленных самок высока, возможно распространение и самцов-волокит. Они не добьются успеха при попытках спариться со скромными самками, но поскольку они быстро оставляют самок, которые не спариваются немедленно, волокиты немного теряют от таких встреч. А спариваясь с легкомысленной самкой, волокиты преуспевают. Они ничего не тратят ни на ухаживание, ни на заботу о потомстве. Партнер волокиты, - легкомысленная самка, - вынуждена нести все затраты при выращивании потомства. Эта модель стабильна в том, что она не эволюционирует к крайнему состоянию, но она не стабильна в том, что не устанавливает конкретное сочетание стратегий самцов и самок; вместо этого она колеблется между альтернативными сочетаниями. Основываясь на эволюционной теории, можно было бы ожидать, что альтернативные стратегии самок обнаруживаются среди особей одного вида, как предсказывает модель Доукинса (Dawkins, 1976), однако оказывается, что они распространены не очень широко. Возможно, эти модели слишком просты или в них нет основных составляющих, о существовании которых мы и не подозреваем. Однако известно несколько примеров альтернативных стратегий самок.
У африканских страусов одни самки насиживают яйца, а другие не насиживают. Самец страуса удерживает большую территорию и строит что-то вроде гнезда на земле. Он спаривается с самкой, которая начинает откладывать в это гнездо яйца. В течение нескольких дней другие самки, которые могут спариваться, а могут и не спариваться с хозяином гнезда, начинают откладывать яйца в то же самое гнездо. Однако они не насиживают и не охраняют яйца, оставляя их на самца или его супругу. В конечном счете в гнезде может оказаться от 30 до 40 яиц, что слишком много для насиживания одной птицей. Насиживающая самка отодвигает некоторые яйца к краю гнезда, и они обычно гибнут, перегревшись на солнце, или их утаскивают хищники.
Бертран (Bertran, 1979) обнаружил, что самка страуса узнает собственные яйца и старается вытолкнуть из гнезда яйца других самок. Она обычно насиживает все собственные яйца и около половины яиц других самок. Насиживающая самка позволяет другим самкам откладывать яйца в ее гнездо, не пытаясь отогнать их. Очевидно, другие самки не связаны с ней тесным родством, так что ее терпимость нельзя объяснить отбором родичей. Самке, по-видимому, выгодно, чтобы ее кладка была окружена чужими яйцами. Хищники, совершающие набеги на гнездо (обычно шакалы и грифы), забирают лишь несколько яиц, так как каждое из них очень велико. Вероятность того, что яйца хозяйки будут похищены, снижается из-за наличия других яиц, в частности из-за того, что ее яйца собраны в середине гнезда. То же самое справедливо и в отношении самки страуса, окруженной выводком: возможно, она вынуждена заботиться о нескольких птенцах других самок, поскольку их присутствие уменьшает вероятность того, что ее собственных птенцов съедят хищники.
Альтернативная стратегия для самки страуса - сделать так, чтобы ее яйца насиживала другая самка, даже если шансы на их выживание невелики. Если откладывать большое число яиц в разные гнезда, то, возможно, такая стратегия и окупится, но она, очевидно, не является предпочитаемой стратегией, так как самки-паразиты часто остаются без самца или их потомство поедают хищники.
Самцы обычно конкурируют за возможность спариться с самками. Такая конкуренция может принимать разные формы, причем большей частью это, очевидно, агрессивное соперничество. Если самец может препятствовать доступу других самцов к самкам, то ему, должно быть, нетрудно заставить самку спариваться с ним, так как у нее не остается другого выбора. У полигинных видов, таких, как благородный олень и морской слон, доминантные самцы могут собрать гарем, который они охраняют от других самцов. Агрессивное соперничество - дело рискованное, сопровождающееся ранениями или даже гибелью, поэтому в царстве животных существует много других форм соперничества самцов.
В случаях, когда самец получает доступ к самкам, прямо защищая их от соперников, проявляется тенденция к социальной организации вида, основанной на гаремной структуре. Создание гарема значительно облегчается, если самки стремятся собраться в группы, что служит защитой от хищников или облегчает использование пищевых ресурсов. У морских слонов образованию гаремов способствует то, что на берегу имеется немного мест, где животные могут спокойно отдыхать. Антилопы и олени образуют стада, чтобы защищаться от хищников. Самцы - владельцы гаремов - часто держатся с краю, защищая самок от соперников, и даже небольшое стремление самок рассеяться чрезвычайно осложнило бы жизнь стада.
У некоторых видов самцы конкурируют не за доступ к самкам, а за оплодотворение. Иногда это называют конкуренцией спермиев. Например, самцы навозной мухи конкурируют за самок, и иногда одному из них удается прогнать другого во время копуляции и занять его место. Паркер (Parker, 1978) показал, что, когда два самца спариваются с одной самкой, сперма второго самца оплодотворяет большую часть яиц. Паркер облучал самцов, что предотвращало развитие яиц в случае оплодотворения их спермиями этих самцов. Если облученный самец копулировал с самкой после того, как с ней спарился необлученный самец, то у самки развивалось только 20% яиц, а если до него - 80% яиц. Каким-то образом спермии второго самца вытесняют большинство спермиев первого самца.
Для копуляционного поведения грызунов характерны многократные интромиссии и эякуляции. Эти действия, По-видимому, облегчают продвижение спермы по половым путям самки. Период покоя, следующий за эякуляцией, очевидно, необходим для завершения продвижения спермиев и для успешной имплантации оплодотворенного яйца в стенку матки (Adler, 1969). Если в этот период с самкой удается копулировать другому самцу, то продвижение спермиев первого самца может быть нарушено и отцом потомства становится второй самец.
Неудивительно, что у видов, у которых возможна конкуренция спермиев, самцы принимают определенные меры предосторожности. Многие млекопитающие совокупляются в уединении, избегая возможного вмешательства. Самец навозной мухи после копуляции сидит на спине самки и охраняет ее, пока та не отложит яйца. Некоторые насекомые после копуляции замазывают половое отверстие самки. Считают, что такие копуляционные пробки предотвращают последующее совокупление у некоторых водяных жуков, дневных и ночных бабочек (Wilson, 1975). Эффективной копуляционной пробкой может служить продолжительное совокупление. Самец комнатной мухи остается в положении копуляции около часа, хотя большая часть спермы передается им самке в течение первых 15 мин. Некоторые ночные бабочки копулируют целый день. У мухи Johannseniella nitida самка съедает самца сразу после копуляции, оставляя только его гениталии, которые служат копуляционной пробкой (Wilson, 1975).
Если самец не в состоянии добиться самки в соперничестве с другими самцами, то он может копулировать украдкой или обманом. Например, самцы лягушки-быка (Rana catesbeiana)конкурируют за территорию в прудах, куда самки приходят откладывать икру. Самки предпочитают определенные участки прудов, обычно те, где вода теплая, а растительность не слишком густая. В таких условиях икринки быстро развиваются, образуя плотные шары, относительно защищенные от нападения пиявок (Howard, 1978). Самцы используют голос и физическую силу, сражаясь за лучшую территорию. Самцы, которые старше и крупнее, обычно побеждают, а небольшие молодые самцы могут остаться вообще без территории. Эти молодые самцы пользуются такой стратегией - они молча сидят около «поющего» самца и пытаются перехватить самок, которых он привлекает. Точно так же молодые морские слоны нередко пытаются присоединиться к гарему самца-доминанта, при этом ведут они себя как самки. Затем они украдкой совокупляются, пока хозяин гарема разбирается с соперником (Le Boeuf, 1974). Молодые самцы благородного оленя также могут пытаться украдкой спариваться с самками из гарема доминирующего самца.
Альтернативные стратегии внутри вида могут использоваться самцами на разных стадиях развития, как в примерах, обсуждавшихся здесь. У некоторых видов отдельные самцы могут использовать обе стратегии.
Система размножения и социальная структура животных
Успешная половая стратегия приводит к развитию определенной системы размножения, которая оказывает глубокое влияние на социальную организацию вида. Так, самец может максимизировать свою приспособленность, спариваясь с большим числом самок и становясь отцом большого числа потомков. Такая полигиния - хорошая стратегия при условии, что эти потомки доживут до половозрелости. Их выживание зависит от экологического окружения и от уровня затрат со стороны родителей, как самца, так и самки. У преуспевающего полигинного самца нет времени на заботу о своих потомках. Либо они должны рождаться самостоятельными, либо самка должна быть в состоянии заботиться о них без его помощи. Полигиния обычно приводит к появлению нескольких преуспевающих самцов и большого числа самцов, не добившихся успеха. В этом случае гарем становится основным признаком социальной структуры, как у многих антилоп и оленей. Самцы соперничают за обладание самками, а молодые самцы вынуждены ждать удобного случая, чтобы спариться украдкой или вызвать доминантного самца на поединок.
Хотя полигиния необычна для пернатых, она часто наблюдается у выводковых птиц, таких, как фазаны, птенцы которых нуждаются в относительно небольшой заботе со стороны родителей. Птенцовую молодь приходится выкармливать, и для успешного выживания птенцов обычно требуется участие обоих родителей. Полиандрия, при которой одна самка спаривается с двумя или более самцами, а самцы - только с одной самкой (Jenni, Collier, 1972). Самки у них заметнее, склонны к территориальному поведению и доминированию в отличие от самцов. После спаривания самка откладывает яйца, которые насиживает исключительно самец. А самка в это время пытается найти другого самца, чтобы отложить следующую кладку.
В общем, образование парного союза связано с заботой о потомстве. У видов, характеризующихся промискуитетом, при котором как самцы, так и самки спариваются более чем с одним представителем противоположного пола, парного союза как такового нет и забота о потомстве минимальна. У других видов система размножения связана частично с условиями обитания, а частично с потребностями молодняка. Это хорошо видно при сравнении разных видов приматов.
Моногамия не типична для млекопитающих, однако она встречается у 14 или несколько большего числа видов приматов (Clutton-Brock, Harvey, 1977). У большинства приматов самцы почти не проявляют заботу о потомстве, хотя защищают семейную группу в случае опасности. У моногамных видов самцы более активно заботятся о потомстве (Passingham, 1982). Например, у мармозеток(Collithrix) и львиных игрунок (Leontideus) обычно рождается двойня. Самец носит их на себе и возвращает матери только для кормления. Точно так же самец тити проводит с малышами больше времени, чем мать. У человека беспомощный младенец обременяет мать, поскольку не может держаться на ней собственными силами. Ребенок, начинающий ходить, не поспевает за взрослыми, и мать вынуждена носить его. В примитивных сообществах эта зависимость может ограничивать частоту рождения детей одной женщиной, так как на ней лежит обязанность добывать пищу для семьи, о чем говорилось выше.
Социальная организация приматов
У приматов велико разнообразие типов социальной организации, что отражает разнообразие их образа жизни и экологии. У многих видов полуобезьян, в том числе у лемуров, лори и долгопятов, источники пищи очень рассредоточены. Живут эти обезьяны на деревьях, питаются насекомыми, и активность у них большей частью ночная. Образ жизни у них одиночный, а социальные контакты осуществляются лишь во время ухаживания и спаривания.
Моногамные виды, такие, как белорукий гиббон (Hylobates lar ), защищают территорию, как большинство моногамных птиц. Детеныши окружены большой заботой и остаются с родителями до половозрелости.
Социальная организация гелад (Theropithecus gelada) очень похожа на то, что мы видим у некоторых антилоп, и, по-видимому, зависит от значительных сезонных изменений имеющихся пищевых запасов. Например, антилопы пала (Aepycerus melampus) образуют стада холостяков из одиноких самцов и гаремные стада из самок под контролем одного самца. Гелады живут на горных лугах Эфиопии. Во время сухого сезона они образуют стаи определенного типа - гаремные, группы только из самцов и группы из молодых. Группы из самцов распространены наиболее широко, а это уменьшает конкуренцию за пищу между группами. В дождливый сезон, когда пищевые ресурсы обильны, эти группы собираются в большие стада, внутри которых гаремы сохраняют неприкосновенность.
Приматы, обитающие в местах с более стабильными пищевыми ресурсами, образуют группы с несколькими самцами. Например, восточная горная горилла (Gorilla gorilla beringei), питающаяся листьями и стеблями лесных растений, имеет небольшие индивидуальные участки. Эти животные держатся маленькими группами, во главе каждой из которых стоит самец с серебристой полосой на спине, но в группу входят и другие самцы. Они не защищают свои участки, которые могут перекрываться с участками других групп.
Павианы, обитающие в саванне, такие, как павиан анубис (Papio anubis), обычно живут стадами, образованными самцами и самками. Доминантные самцы имеют преимущественное право доступа к самкам, пище и воде. Они объединяются против соперников для удержания своего социального положения и против других особей стада при защите кормящих матерей и детенышей. Доминантные самцы также объединяются при защите против хищников. Богатые местообитания обычно привлекают большое количество хищников, и объединение самцов является значительным преимуществом в таких условиях.
Результаты сравнительных исследований (например, Brown, 1975) позволяют предположить, что в суровых условиях стадо павианов возглавляет один самец, тогда как в богатых местообитаниях в каждом стаде имеется группа объединившихся доминантных самцов. Плащеносные павианы гамадрилы (Papio hamadryas) из Эфиопии типичны для первой группы, тогда как желтый павиан (P. cynocephalus) представляет другую крайность. Павианы анубис (P. anubis) и медвежий (P. ursinus) вэтом отношении образуют промежуточную ступень. Павианы гамадрилы подвергаются сильному половому отбору, и доминантных самцов легко отличить по большой гриве. Соперничество самцов гораздо менее интенсивно у желтых павианов, и у них в гораздо меньшей степени выражен половой диморфизм.
Сотрудничество является отличительной чертой поведения человека. Первобытные люди, очевидно, охотились вместе и делили добычу на всех (Isaac, 1978). Как и у других приматов, у человека существует разделение труда между полами. В сообществах шимпанзе охотятся самцы, тогда как самки гораздо чаще, чем самцы, собирают термитов (McGrew et al., 1979). В племенах туземцев, живущих и в наши дни охотой и собирательством, мужчины занимаются в основном охотой, тогда как женщины собирают растительную пищу (Lee, DeVore, 1968). Новорожденные у человека крайне беспомощны, так что женщины часто ограничены в своей трудоспособности. Человек не единственный вид с таким типом социальной организации. Самки гиеновой собаки(Lycaon pictus) некоторое время держатся в норах со щенками и целиком зависят в это время от совместной охоты самцов и от того количества мяса, которое принесено домой для дележки.
Человек уникален среди млекопитающих разделением труда внутри полов (Passingham, 1982). Различные индивидуумы играют разную роль, значительно увеличивая, таким образом, эффективность кооперации. Например, пигмеи племени мбути объединяются для охоты на слонов. Один человек останавливает слона спереди, в то время как второй сзади перерезает сухожилие на одной ноге, а затем на другой (Turnbull, 1966). Такие охотники общаются при помощи жестов, чтобы остаться незамеченными жертвой. Таким же образом первобытные люди могли достичь сложной кооперации, не пользуясь языком.
Биологические механизмы человеческой сексуальности.
Сексуальность человека обусловливается сложным взаимодействием биологических факторов - анатомии и физиологии, нервной и гуморальной систем, сексуальных реакций и переживаний.
В процессе филогенеза и онтогенеза сексуальность человека освобождается от биологической детерминированности и больше зависит от психологических и социокультурных факторов. Так, оргазм характерен только для человека и не встречается у животных.
Центральная нервная система, и прежде всего кора головного мозга, играет огромную роль в половой жизни человека. В свою очередь, она неразрывно связана с эндокринной системой (происходит нейросекреция - выделение нейромедиаторов, передача нервных раздражителей, стимулирующих железистые клетки эндокринной системы).
Основное влияние на сексуальную реактивность оказывает кора головного мозга. Биоэлектрическая активность коры при оргазме сходна с таковой при эпиприпадке. Повреждение коры может привести к ослаблению или исчезновению сексуальной активности, гиперсексуальности, девиантному поведению либо к нарушению полового созревания. Кора головного мозга представляет собой первый уровень регуляции сексуальных функций, связанный с условными рефлексами (сигналом является изменение внешней среды в связи с возникновением или исчезновением сексуального напряжения), сферой представлений и воображения, элементарными ассоциациями. Сексуальный партнер как сексуальный раздражитель воздействует на другого партнера через кору головного мозга посредством органов чувств, вследствие чего возникает возбуждение и стремление к сексуальному контакту с партнером. Таким образом, выбор партнера является результатом предыдущих контактов человека с другими людьми.
Важным аспектом зрелого сознания является половое, становление которого представляет одну из важных сторон формирования личности. Поэтому при негативном сексуальном воспитании эмоциональное напряжение при действии раздражителя может восприниматься отрицательно и в дальнейшем превращаться в “антисексуальное” напряжение, требующее разрядки несексуальным способом. Как избыток, так и недостаток раздражителей нарушает функционирование коры головного мозга.
Центром влечений, ответственным за эмоции и управляющим инстинктами самосохранения и сохранения вида, является гипоталамус - подкорковое образование мозга. В передней части гипоталамуса находится центр, высвобождающий сексуальную активность, а в средней и задних долях - центр (гормональный сексуальный центр) обусловливающий сексуальное влечение опосредованно через секрецию гонадолиберина, которая активизируется норадреналином и тормозится серотонином. Гонадолиберин стимулирует продукцию гонадотропных гормонов гипофиза - лютеинизирующего гормона (ЛГ) и фоликулостимулирующего гормона (ФСГ), которые влияют на половые железы. У мужчин гипоталамус стимулирует равномерное выделение ЛГ и ФСГ, а у женщин – циклическое (вначале преобладает ФСГ, затем ЛГ).
Сексуальные центры гипоталамуса оказывают регулирующее влияние на сексуальные центры спинного мозга, в свою очередь, подвергаясь регулирующим влияниям лимбической системы (тормозящее влияние) и коры головного мозга.
Спинномозговые центры отвечают за реакции в половых органах (за эрекцию отвечает крестцовый отдел, за эякуляцию, а также за секрецию половых желез — поясничный отдел). Там же происходит переключение как простых рефлексов, так и рефлексов с участием высших отделов нервной системы. Поражение спинномозговых центров или проводящих путей, по которым импульсы передаются к половым органам, приводит к выпадению соответствующей сексуальной реакции. При нарушении проведения импульсов от коры головного мозга к спинномозговым центрам наблюдается подобный, но менее выраженный эффект. Поражение путей, передающих импульсы от половых органов в мозг, вызывает снижение чувства сексуального наслаждения иногда вплоть до отсутствия оргазма (при невысокой возбудимости).
Сложные формы инстинктивного поведения.
Поведение животных вовсе не исчерпывается простыми видами реакций; у животных, обладающих центральной нервной системой (построенной по типу цепочки нервных ганглиев) можно наблюдать и гораздо более сложные формы поведения, которые вызываются относительно элементарными сигналами, но развертываются в виде сложных программ, которые приводят к высокой степени целесообразным дифференцированным актам. Интерес этих сложных форм поведения заключается не только в их высокой целесообразности, но и в том, что эти программы наследственно закреплены. Иногда эти наследственные программы видового поведения являются настолько сложными, что создают впечатление разумных видов приспособления животных к окружающей среде; поэтому у некоторых исследователей возникла тенденция толковать их как разумные интеллектуальные формы поведения животных и приписывать таким животным разум, близкий к человеческому.
На этих формах поведения, которые называются инстинктами и которые в истории науки всегда вызывали массу трудностей и массу споров, мы сегодня и остановимся.
Под инстинктом принято понимать «целесообразное поведение при отсутствии сознания цели». Иначе говоря, речь идет здесь о тех сложных наследственно программированных формах целесообразного поведения, посредством которых животное без специального обучения начинает приспосабливаться к условиям окружающей среды. Эти формы поведения преобладают на ранних этапах развития беспозвоночных, например у насекомых эти формы поведения являются основными, и иногда почти что единственными; у других животных – низших позвоночных – остаются преобладающими и только постепенно уступают свое место более сложным индивидуально изменчивым формам поведения, вырабатываемым в процессе прижизненного обучения.
Формы поведения простых дождевых червей, например, активное закапывание в землю или активные движения в определенную сторону можно было наблюдать у любого дождевого червя с сохранным передним ганглием.
Но у дождевого червя можно наблюдать и гораздо более сложные формы поведения, объяснение которых является гораздо более трудным.
Для того, чтобы сохранить пищу на зиму и закрыть вход в свою нору, дождевой червь очень часто втаскивает в свою нору листья, которые остаются там и которыми червь может питаться в течении довольно длительного периода. Исследователи обратили внимание на удивительный факт: втаскивая в нору лист, червь всегда берет его за передний конец; казалось бы во всех возможных способах он выбирает наиболее целесообразный способ введения листа в узкое отверстие норы. Казалось бы, червь различает форму листа и использует его острый конец, позволяющий ему провести нужную операцию самым экономным путем. Однако, как расшифровать те механизмы, которые лежат в основе таких форм целесообразного действия.
Ученые провели ряд исследований, которые показали, что целесообразное поведение червя, втаскивающего лист в нору за узкий конец, на самом деле гораздо проще, чем это кажется, и что оно детерминировано гораздо более элементарными и вполне понятными для нас факторами.
Прежде всего – нужно ответить на вопрос: существует ли у червя восприятие формы? Один из исследователей провел очень остроумный опыт: он взял лист и вырезал из него кусочек, по форме напоминающий лист, только так, что в этом кусочке острый конец был обращен вниз, а тупой конец – вверх. В результате червь всегда втаскивал в нору вырезанный из листа кусочек за тупой конец, а не за острый. Значит, в основе действия червя лежало не восприятие форм листа, а какой – то другой фактор.
может быть в основе этих действий червя лежат какие-то химические сигналы? У червя хорошо развитые химические рецепторы, расположенные на переднем конце его тела. Чтобы проверить, была проведена серия опытов; она принадлежит исследователю Мангольду. Этот исследователь взял палочки одинаковой толщины и смазал один конец эмульсией, полученной из верхней (острой) части листа, а другой конец – эмульсией, полученной из нижней (тупой) части листа. Оказалось, что в этом случае червь брал эти палочки всегда за тот конец, который был смазан эмульсией из острой части листа. Значит, в острой части листа были какие-то химические вещества, которые являлись сигналом для червя к тому, чтобы брать лист за острый конец. Если в контрольных опытах оба конца палочек смазывались нейтральной желатиной, такая избирательность исчезала, и червь втаскивал палочки в нору то за один, то за другой конец.
Следовательно, можно предполагать, что целесообразное поведение червей вовсе не является реакцией на форму листа, а реакцией на химические вещества, которые отличают вершину листа от его основания. Таким образом, это поведение определяется гораздо более простыми факторами и прежде всего химическим чувством, и сложное поведение возникает как результат относительно простых сигналов.
Если у червя, описанные формы врожденного поведения, являются еще относительно простыми, то у высших беспозвоночных, например, насекомых – наследственно программированные формы поведения приобретают несравненно более сложный характер и становятся действительно удивительными по своей сложности и дифференцированности. Эти формы поведения являются ведущими формами у беспозвоночных; они могут служить наиболее яркими примерами того, что обычно называется «инстинктивным» поведением.
В чем заключается основной принцип организаций этих сложных форм поведения, достигающих предельного развития у насекомых? Этот принцип заключается в следующем: в процессе эволюции какими-то еще неизвестными путями (может быть, путем мутации, может быть, иными путями) создаются как более соответствующие условиям существования этих животных – форма строения их тела и форма их поведения.
Крыло мухи или бабочки является идеальным прибором для полета. Хоботок пчелы является идеальным прибором для того, чтобы добывать нектар цветов; как известно длина хоботка такова, что он приспособлен как раз для добывания нектара из всех растений, которыми питаются пчелы. Строение тела у любого насекомого поражает своей целесообразностью и приспособленностью к условиям его существования. Такое целесообразное строение тела результат законов эволюции и отбора, изученных Дарвиным. Такие же целесообразные и приспособленные к условиям среды характером отличается не только строение тела беспозвоночных, но и программа их поведения.
Сложные наследственно заложенные формы поведения, одинаковые у всех представителей данного вида, с которыми животные рождаются, но которые являются очень сложными, а иногда производят впечатление разумных, и называются инстинктами.
Известно, что комар, когда приходит время кладет яичко на поверхность воды. Если бы он отложил яички не на поверхность воды, а на сухом месте, то яички погибли бы. Комар избирает определенный способ откладывания яичек на поверхности воды, являющейся наиболее целесообразным, и делает это без всякого специального научения. Личинки многих жуков делают шелковый кокон во много раз больше своего собственного размера, причем величина этого кокона соответствует той, которая понадобится для зрелого жука, который из него выйдет.
Оса проделывает удивительную технику откладывания яичек. Есть виды, которые откладывают свои яички в теле гусеницы. Для того, чтобы эти яички сохранились длительное время и чтобы личинки, которые вылупятся из них, имели бы возможность питаться, оса производит удивительную операцию. Она забирается на гусеницу и жалом укалывает ее в двигательные ганглии. Гусеница не умирает, но обездвиживается, и, когда из яичек вылупляются личинки оси, у них есть своя свежая пища – тело гусеницы, мясо которой не разложилось, но которая остается обездвиженной и, таким образом, дает возможность питаться.
Вся организация такого сложного поведения поражает своей целесообразностью. Можно подумать, что оса может делать расчеты, показывающие, где у гусеницы расположены двигательные ганглии. Можно подумать, что пчела, которая строит соты по максимально экономному плану (а исследователи-геометры подсчитали, что нельзя придумать более экономной формы построения сот из воска, чем многоугольная форма ячеек), делает это по каким – то расчетам. То же самое можно подумать об одной из личинок, которая разрезает листья и свертывает трубочки из них, проделывая это с удивительным геометрическим «расчетом» и разрезает лист так, как может его разрезать только математик, который с предельной точностью расчитывает наиболее экономную форму разреза листа.
Наиболее интересным является, однако, тот факт, что во всех этих случаях это поведение одинаково у всех особей данного вида; каждая особь не должна вырабатывать свою программу поведения в индивидуальном опыте, и одновременно это поведение оказывается целесообразным, соответствуя тем условиям, среди которых живет данное животное.
Известно, что личинка европейского угря в Балтийском и Северном морях совершают путешествие через всю Атлантику до Бермудских островов в Северной Америке, и только там, после того, как угорь окрепнет, он возвращается снова на родину. Это путешествие занимает три года, и личинки в один миллиметр диной покрывают расстояние в тысячи километров, каждый раз приходя в одно и тоже место, а затем возвращаясь обратно. В то же время личинки американского угря делают аналогичное путешествие в обратном направлении. И здесь сложное поведение оказывается точно программированным, хотя те ориентиры, по которым действует животное, остаются неизвестными. Аналогичное можно наблюдать, изучая перелеты птиц, в которых самое интересное заключается в том, что птицы делают перелеты, занимающие много и много тысяч километров, точно ориентируясь в направлении своего полета и точно возвращаясь к соответствующему сезону на исходное место.
Все это показывает, что многие формы поведения насекомых и низших позвоночных состоят из сложнейших врожденных программ, одинаковых для всех представителей данного вида и предельно целесообразных в обычных для животных условиях существования. Именно это дало возможность некоторым авторам определить инстинкт, как «целесообразное поведение без сознания цели» и указать на четыре основных качества такого инстинкта; наследственность и независимость от обучения, однородность, одинаковость у всех особей данного вида и приспособленность к условиям существования.
Теперь возникает самая большая проблема: каким образом формируются эти сложнейшие формы поведения? Что лежит в их основе, что их вызывает? Чем они отличаются от подлинных разумных форм поведения? И, наконец, как эти врожденные формы поведения относятся к индивидуально изменчивым формам поведения животных? Многие проблемы из организации инстинктивных программ поведения (например, перелет птиц) до сих пор остаются не раскрытыми в науке.
Сложные виды инстинктов возникают, конечно, не у медузы или морской звезды; их еще нет у животных с сетевидной нервной системой; они еще только намечаются у животных с элементарными формами ганглионарной нервной системой (например, у червей). Лишь сложно построенная ганглионарная нервная система является аппаратом, который позволит осуществлять описанные выше наследственно программированные формы поведения.
Рассмотрим некоторые данные о строении ганглионарной нервной системы насекомых (пчелы, мухи, бабочки, жуки).
Известно, что нервная система пчелы состоит из цепочки нервных узлов, расположенных по всей длине ее тела. Однако, эти узлы неодинаковые; узлы, расположенные на головном конце тела и тесно связанные с периферийными экстерорецепторами – имеют гораздо более крупные размеры и гораздо более сложное строение.
Известно, что на переднем головном конце тела пчелы расположены рецепторы, значительно более дифференцированные и сложные, чем те рецепторы, которыми располагает червь. У пчелы есть все те органы чувств, которые есть у человека, а кроме того, есть и некоторые органы чувств, которых нет у человека. На передней части головы расположены обонятельные органы пчелы, включенные в усики, причем эти усики пчелы являются органами двойного значения: это орган для ощупывания и орган для обоняния. Комбинация этих двух рецепторов в органе дает возможность говорить о том, что пчела имеет своеобразные «топохимические» органы, которые отражают как форму предмета, так и его запах. Пчела обладает тончайшим обонянием, которое позволяет ей различать запах на длинном расстоянии. Тонкость обоняния пчелы проявляется, например, в том, что она может различать запах померанцевого масла, выделяя его из 140 других запахов.
У пчелы есть орган вкуса, причем вкусовые рецепторы расположены в хоботке, во рту, в ножках. Тонкость вкусовых ощущений меняется у пчелы в зависимости от ее состояния. Голодная пчела или бабочка различает очень тонкую концентрацию раствора сахара, а сытая пчела различает только более концентрированный.
У пчелы есть слуховые и вибрационные органы, воспринимающие колебания большого диапазона частот, причем эти органы слуха и вибрации расположены в перепонках, которые натянуты на ножки. Возможно, что насекомые различают гораздо более тонкие колебания звуковых волн, чем человек. Человек различает колебания частотой от 20 до 20 тысяч в секунду, а насекомые, по всем данным, реагируют и на ультразвуковые колебания. Возможно, что восприятие ультразвука этими тимпанальными перепончатыми органами создает у насекомых возможность локации.
У насекомых есть, наконец, и в высшей степени развитое зрение. Глаза насекомых, в отличии от глаз позвоночных, состоят из многих тысяч отдельных ячеек, каждая из которых свою световую точку, разлагая образ на массу отдельных элементов так.
Возбуждения, возникающие в этих органах, передаются затем в передний ганглий, который у червя носил еще скромный характер, у всех насекомых сильно развивается и обнаруживает уже очень сложное строение.
В этом переднем ганглии можно выделить три совершенно разные части: передняя часть, куда доходят волокна от зрительного рецептора; средняя часть, куда доходят волокна от обонятельного рецептора; задняя часть, куда доходят волокна из ротовой области. Таким образом, у пчелы есть дифференцированный передний ганглий, который во многих отношениях можно сравнить с передним мозгом позвоночных.
Значительную новизну представляет и гистологическое строение этого ганглия. Русский гистолог Заварзин показал, что передний ганглий у пчел построен совершенно не так, как у червей. Он построен не по принципу хаотичного скопления нейронов, а по принципу плоскостного расположения нейронов, или по «экранному» принципу. Это строение позволяет пчеле организовано отражать световые сигналы, доходящих из среды, и воспринимать ту конфигурацию, в которой расположены эти раздражители. Наоборот, в отличие от этого, строение обонятельной части переднего ганглия не имеет такого «экранного» строения и носит характер клубочков, которые расположены в беспорядке. Следовательно пчела имеет возможность отражать зрительные раздражения от «экрана», но обонятельные раздражения – более диффузно.
Этот передний ганглий пчелы и производит синтез обонятельных, зрительных и вкусовых раздражений; по всей вероятности, этот передний ганглий насекомого и служит аппаратом, который позволяет выработать сложные программы поведения. Эти сложные программы, вырабатываемые в переднем ганглии, переводятся на следующий грудной ганглий, который дает возможность перешифровать складывающие системы на двигательную программу и пустить в ход сложные двигательные программы поведения.
Характерно, что эти сложные программы поведения возникают в ответ на сигналы, полученные из среды, не вырабатываются каждым насекомым в процессе его жизненного развития, а в готовом виде существуют у всех представителей данного вида. Иначе говоря они закодировано наследственно, и доходящий до переднего ганглия раздражитель лишь возбуждает эти готовые программы, приводит их к реализации.
Вот то немногое, что известно о механизмах, лежащих в основе описанных сложнейших форм инстинктивного поведения высших беспозвоночных.
Возникает основной решающий вопрос: чем вызываются и как протекают эти инстинктивные формы поведения. Возникают ли эти сложные программы в результате детального анализа ситуации или же в результате относительно простых стимулов, которые пускают вход программы, заложенные в нервных ганглиях, и ведут к дальнейшему развертыванию врожденных форм поведения? Если инстинктивные формы поведения возникают в результате сложного анализа окружающей обстановки и соответствующей синтетической работы – они приближаются к разумным формам психической деятельности. Если же инстинктивные формы поведения осуществляются по наследственно закрепленным программам, но вызываются к жизни относительно простыми сигналами, которые служат лишь толчком к реализации сложных программ, то такое поведение по своему типу приближается к рефлекторному, и разница заключается только в том, что в простых рефлексах ответы носят относительно простой характер (выделение слюны, вздрагивание мышцы). А в интересующем нас случае ответом является целая программа, которая включается тогда, когда появляется известный сигнал. Наибольший интерес представляют исследования ряда немецких и английских исследователей; Лоренца, Тинбергена, Торпа и других, объединенных в целую школу, которая получила название «этология» («Этос» – действие, поведение, следовательно, этология – наука о поведении). В отличии от бихевиоризма – американская наука о поведении, этология занимается не только анализом индивидуально формируемых видов поведения (навыков), но главным образом, анализом механизма врожденных форм поведения и механизмов, которыми эти врожденные формы поведения вызываются к жизни.
Основная идея этологии заключается в том, что инстинктивное поведение вовсе не заключается в спонтанно возникающих программах, но в том, что эти программы возникают в ответ на определенный набор внешних раздражителей, которые можно изучить. Значит, к инстинктам можно подходить как к сложным детерминированным рефлекторным актам, которыми животное отвечает на появление вполне определенных внешних сигналов.
Комар откладывает яички на поверхность воды. Это унаследованная и в высокой степени целесообразная программа поведения. Естественно, что это поведение возникает тогда, когда наступает пора откладывать яички и когда создается биологическая необходимость этого действия, но, как показали опыты, для откладывания яичек необходим специальный сигнал и таким сигналом является блеск воды. Когда самка, которая должна откладывать яички, раздражается блеском воды, то она откладывает яички на блестящую поверхность. Было установлено, что достаточно показать комару зеркало или даже блестящее стекло, чтобы комар отложил яички на этой поверхности.
Второй пример. Паук, который сидит в паутине набрасывается на муху, запутавшуюся в паутине, и пожирает ее. Это тоже врожденная программа поведения. Сигналом, вызывающим реализацию этой программы действия, является вибрация. Характерно, что достаточно приложить к паутине вибрирующий камертон, чтобы вызвать такое же поведение: паук вылезает из своего гнезда и бросается на камертон, производя все движения, которые он производил в отношении мухи.
Аналогичные факты можно наблюдать и в поведении позвоночных. Лягушка подстерегает свою жертву; когда она видит летящую мошку, она делает прыжок и заглатывает жертву. Таким образом, вид летящей мошки вызывает у лягушки инстинктивные действия. Если на маленьком волоске прикрепить бумажку и сделать так, чтобы это бумажка мелькала перед лягушкой, последняя непременно бросится на нее. Значит, у комара инстинктивное действие, вызывается блеском, а у паука – вибрацией, у лягушки – мельканием.
Профессор Анохин П. К. и его сотрудники показали, что когда в гнезде грача появляются птенцы и отец подлетает к гнезду с червяками, которые он несет в клюве, возникает следующее поведение птенцов, как только отец подлетает к гнезду, птенцы сейчас же раскрывают рты и готовятся к тому, чтобы заглотать пищу. Чем же вызывается это сложное инстинктивное действие? Этим сигналом является звук «карр», который испускает подлетающий грач, и колебание, которое возникает от того, что он садится на край гнезда. И если в искусственных условиях давали птенцам этот звук «карр» или колебали край гнезда, грачата делали точно такие же движения, какие они делали в ожидании пищи.
Маленький только что родившийся щенок ищет сосок матери и как только находит, начинает энергично сосать. То же самое делает и только что родившейся ягненок. Это показывает, что у этих только что родившихся детенышей, которых никто еще не учил, есть целесообразные программы инстинктивного действия. Очень интересные исследования были проведены одним из советских авторов, профессором Слонимом. Опыты ставились так: вся ситуация – щенок под телом матери – разлагалась на отдельные компоненты. Щенку дали гладкую теплую грелку, она не вызывала сосание; но если ему давали теплую шерсть – начинались сосательные движения. Значит сигналом, который пускает в ход эту программу действия, является шерсть. Сигнал, вызывающий сосательные движения у ягненка, может показаться неожиданным: таким сигналом является затемнение темени ягненка. Этот удивительный факт объясняется тем, что только что родившийся ягненок начинает сосать, когда он подходит под мать и когда его головка затемняется.
Из всего сказанного можно сделать два очень существенных вывода. Первый вывод заключается в том, что удивительные по своей целесообразности инстинктивные врожденные программы действий, которые у беспозвоночных являются основными формами поведения, построены по рефлекторным механизмам. Они вызываются элементарными факторами – относительно простыми сигналами, которые вызывают включение всей сложной программы. Такими факторами является отражение отдельных простейших свойств или признаков (блеск, вибрация, мелькание, колебание, затемнение темени, мягкая шерсть). Значит, врожденные сложные программы поведения включаются простейшими сенсорными факторами.
Второй существенный вывод заключается в том, что программы сложного инстинктивного поведения включаются не любыми свойствами, воспринимаемыми животными, а только теми свойствами, которые соответствуют способу жизни животных или, как говорят, их «экологии». Иначе говоря, сложные врожденные программы поведения вызываются сигналами, которые соответствуют экологическим условиям жизни животных.
Существуют два очень близких вида птиц: домашняя утка и дикая кряква; домашняя утка питается растительной, а кряква – разложившейся животной пищей. Поэтому инстинктивные программы поведения вызываются у утки растительными запахами, а на запах аммиака утка никак не реагирует, а кряква же – наоборот. Те сигналы, которые отражают образ жизни, включают в действие эти врожденные программы поведения. Существенно, что типовые способы существования животных определяют не только то, какие сигналы действуют на животное, но и то какие программы действий включаются этими раздражителями. Вот очень разительный пример: собака и лиса представляют зоологически очень близкие разновидности, но экологически они очень различны: собака живет в домашних условиях и питается готовой пищей, которую ее дает человек, а лиса живет в диких условиях и сама добывает себе пищу. Это формирует противоположное поведение, возникающее у обеих при виде пищи или ощущения запаха пищи. У собаки в этом случае начинает выделяться слюна (вся павловская физиология построена на изучении этого безусловного рефлекса), у лисицы возникает обратная реакция – запах или вид пищи тормозит слюну, потому что ей сначала надо добыть пищу, а выделение слюны может мешать добычи пищи.
Аналогичные факты зависимости избирательного отношения к разным сигналам от условий жизни (экологии животного) можно видеть и при изучении высших беспозвоночных.
Известно, что пчела обладает глазом, который может различать известные признаки формы, но какие из этих признаков реально используются как сигналы, которые вызывают известные инстинктивные программы поведения: подлетание, попытка выпускания хоботка и акт вытягивания сока? Немецкий психолог Фриш провел очень интересные опыты: становилось несколько баночек. В одних был сахарный раствор, в других вода без сахара. Баночки с сахарным раствором покрывались крышками с одним изображением (скажем, треугольником), а баночки, в которых не было сахара, покрывались крышечками с другим изображением, например, квадрата. Таким образом фигура треугольника всегда подкреплялась, а фигура квадрата оставалась без подкрепления. Ставится вопрос – возможно ли вызвать у пчелы различие этих форм? Казалось бы, такие простые геометрические фигуры, как треугольник или квадрат, легко различить, в то время как более сложные фигуры различить труднее. Однако опыт показал обратное. У пчелы было трудно выработать различие простых геометрических фигур, но было очень просто выработать различие простых геометрических фигур (например, вызвать реакцию треугольника и квадрата или круга и квадрата). Более сложные фигуры по своим основным признакам (лучеобразный характер, изрезанность) напоминают цветы, а круг, квадрат и треугольник ничего не напоминают. Поэтому в процессе видового опыта сигналами, которые хорошо улавливают пчела или бабочка, стали какие-то признаки, напоминающие фигуру цветка.
Вслед за Фишем были проведены опыты другим немецким исследователем, Матильдой Герц. Эти опыты показали, что признаком, который хорошо улавливается насекомым, является признак изрезанности, или признак радиального расположения лучей.
Совершенно то же самое выяснилось и при исследовании реакции на цвета. Оказалось, что у пчелы очень трудно выработать различие чистого зеленного от чистого красного цвета или ахроматических цветов, но сравнительно легче выработать различие смешенных цветов (желто-зеленого от желто-оранжевого). Это происходит потому, что цветы всегда являются комплексными по своим цветовым оттенкам и эти комплексные оттенки прочно вошли в экологию пчелы. Поэтому она великолепно различает зелено-желтые цвета от зелено-голубых, красно-фиолетовые от ультрафиолетовых. Дифференцированные реакции на них и является составной частью ее естественного поведения, и этим определяется тонкость различая пчелой комплексных цветовых оттенков.
Все приведенные факты показывают, что сложнейшие инстинктивные программы поведения могут вызываться относительно простыми сигнальными признаками, и что эти врожденные программы вызываются принципиально тем же путем, как и более простые безусловные рефлексы.
Существенным является тот факт, что на этом этапе развития деятельность животных вызывается и регулируется не отражением сложных образов или сложных картин целой ситуации, а отражением элементарных свойств, отдельных изолированных раздражителей, действующих на животное. Вот почему эту стадию развития можно назвать сенсорной психикой, отличая ее от более сложной стадии перцептивной психики, на которой основным фактором, формирующим поведение животного, является отражение комплексных сложных форм или предметов, воспринимаемых животным.
Стадия сенсорной психики и вызываемого отдельными сигналами инстинктивного поведения еще не обеспечивают нужного анализа ситуации, и именно поэтому инстинктивное поведение может легко потерять свою целесообразность.
Есть разновидность осы, которая прежде чем откладывать яички в норе или зарыть там пищу, проделывает сложную и очень целесообразную программу врожденного поведения. Она оставляет добычу на входе, вползает в нору, обследует ее, и только если в норе никого нет, втаскивает в нее пищу, оставляет ее в норе и улетает. Был произведен опыт: когда оса прилетает в нору, оставляет у входа добычу и уходит обслеживать норку, экспериментатор отодвигает эту добычу на два сантиметра. Оса выходит из норы, не находит оставленную добычу на прежнем месте, ползет дальше, находит добычу, снова подтаскивает ее к норе, а затем опять... оставляет у входа и снова ползет в нору. Если в это время экспериментатор опять отодвигает добычу, оса, вернувшись из норы, снова ищет добычу, снова подтаскивает, и уходит обследовать нору. Такое действие продолжается и дальше, в результате оса так и оказывается не в состоянии втащить добычу в нору, подчиняясь косной, твердо упроченной программе врожденного инстинктивного поведения.
Это убедительно показывает, что инстинктивная программа действует очень четко, очень приспособлено в стандартных условиях и оказывается совершенно неадекватной в изменившихся условиях.
Другой пример. Пчела откладывает мед в ячейки, а потом запечатывает эти ячейки. Был проведен опыт. Дно ячеек было срезано, и мед отложенный пчелами, проваливался в пустоту. В этом случае поведение пчелы не менялось. Она откладывала столько меда, сколько обычно откладывалось в ячейку, запечатывала пустую ячейку, а затем закрывала мед в следующую ячейку. Таким образом, инстинктивное поведение пчелы не приспособлялось к изменившимся условиям, и ее инстинкт, оставаясь мало пластичным, легко теряет свой целесообразный характер.
Инстинктивное поведение, осуществляющееся по сложной наследственно упроченной программе, четко приспособлено к стандартным условиям видового опыта, но оказывается неприспособленным к изменившимся индивидуальным условиям. Поэтому достаточно немного изменить стандартные условия, чтобы инстинктивное поведение теряло свой целесообразный характер. Именно этим инстинктивные формы поведения животных отличаются от разумных форм поведения, которые меняются в соответствии с изменившимися условиями.
Классификация форм индивидуально-приспособительной деятельности животных.
Формы обучения животных весьма разнообразны и обычно их подразделяют на три основные категории: неассоциативное обучение, ассоциативное обучение и когнитивные процессы. О. Меннинг (1982) и Д. Дьюсбери включали в свои классификации также и «инсайт-обучение».
Классификация форм обучения и когнитивных процессов составлена на основе классификаций О. Меннинга, Д. Дьюсбери, Р. Томаса, Дж. Пирса и др.:
Неассоциативное обучение: привыкание.
Ассоциативное обучение:
2.1 классические условные рефлексы;
инструментальные условные рефлексы.
3. Когнитивные процессы:
3.1 латентное обучение;
3.2 выбор по образцу;
обучение, основанное на представлениях о:
пространстве;
3.3.2 порядке стимулов;
времени;
3.3.4 числе.
3.4 элементарное мышление.