

FIG.
7-21 All tRNA molecules have a characteristic four- lobe structure
that results from internal base pairing of some of the nucleotides.
Each lobe of the tRNA molecule has a distinct function. Several of
the lobes are characterized by the inclusion of unusual
nucleotides. These nucleotides are formed by enzymatic
modification of the nucleotides directly coded for by the DNA;
that is, the DNA does not have additional nucleotides that directly
call for the insertion of nucleic acid bases other than adenine,
uracil, cytosine, and guanine into the RNA. One of the lobes,
designated the DHU
or D lobe,
contains dihydrouracil (DHU). This lobe binds to the enzyme
involved in forming the peptide during translation. The Ti|fC lobe
contains the sequence ribothymine (T), pseudouracil (i|i), and
cytosine (C). This lobe binds to the ribosome. A third lobe, which
also contains modified purines, is designated the anticodon
lobe
because it is complementary to the region of the mRNA, the codon,
that specifies the amino acid to be incorporated during protein
synthesis. The 3'-OH end always has the terminal sequence ACC,
which is where the amino acid binds. This terminal sequence is
usually referred to as the CCA
end,
reading from the 5'-P end of the tRNA molecule.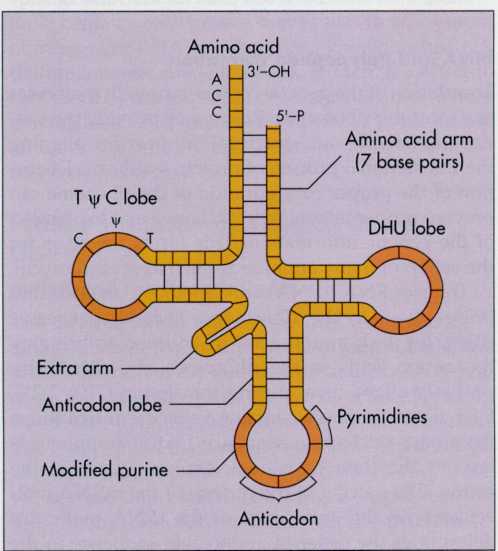
212 Chapter 7 microbial genetics: replication and expression of genetic information
FIG.
7-22 During
protein synthesis the codons of the mRNA are translated into an
amino acid sequence at the ribosome. Each codon of the mRNA matches
an anticodon of a tRNA so that the proper amino acid sequence is
formed. The start codon AUG specifies the insertion of formyl
methionine (f-Met) at the peptidyl site. A second amino acid is
aligned at the aminoacyl site by the pairing of a tRNA with the
codon. Formyl methionine is transferred to the amino acid at
the aminoacyl site with the formation of a peptide bond. The mRNA
then moves along the ribosome so that the tRNA with its two attached
amino acids moves to the peptidyl site. A new amino acid is aligned
at the aminoacyl site, again by pairing of the appropriate tRNA
with the codon. The two amino acids are transferred to the amino
acid with the formation of a new peptide bond so that the peptide
chain now has three amino acids. The process is repeated over and
over to form the long polypeptide chain of amino acids joined by
peptide bonds in the sequence specified by the mRNA.
the
messenger RNA codon. The first and second bases of the codon
sequence are therefore more important in matching the codon
with the anticodon. As a result, a codon may pair with more than one
anticodon that differs only in the third base position, a
phenomenon called wobble.
Transfer
RNA brings amino acids to the ribosomes and properly aligns them
during translation.
Forming
the Polypeptide
During
translation, transfer RNA molecules bring individual amino
acids to be sequentially inserted into the polypeptide chain (FIG.
7-22). The codon of the messenger RNA that specifies where the
synthesis of a polypeptide is initiated is called the start
codon.
The
tRNAs arrive in the order specified by the codons in the mRNA as the
mRNA moves across the surface of a ribosome. When tRNA molecules
arrive at the ribosome, the proper anticodon pairs with its
matching
codon of mRNA. The amino acid is thus aligned so that it can be
covalently bound to a growing peptide chain. After a peptide
bond is established between amino acids already in the polypeptide
chain and the newly aligned amino acid, the messenger RNA then
moves along the ribosome by three nucleotides. The movement of
messenger RNA, transfer RNA, and the growing polypeptide chain along
the ribosome is known as translocation.
The
process is repeated over and over, resulting in the elongation of
the polypeptide chain. Eventually, one of the nonsense codons
appears on the mRNA as it moves across the ribosome. Since no tRNA
molecule pairs with the nonsense codon, the translational process is
terminated and the polypeptide is physically released from the
ribosome.
Translocation
is the movement of messenger RNA, transfer RNA, and the polypeptide
chain along the ribosome.
EXPRESSION
OF GENETIC INFORMATION 213
REGULATION
OF GENE EXPRESSION
Cells
have structural genes that encode the information for specific
polypeptide sequences of proteins. Cells also have regulatory genes
that code for gene expression. It would be inappropriate and energy
depleting for the entire genome to be expressed at one time. By
controlling which genes of the organism are to be translated into
functional enzymes, the cell regulates its metabolic
activities. While some genes are constantly "turned on,"
others are expressed only in response to the immediate needs of the
cell. It is advantageous for a cell to regulate gene expression
so that it can conserve its resources. This is important to conserve
the supply of energy, as well as to utilize sparingly the limited
pool of metabolic intermediates. By regulating gene expression
the organism modifies its phenotype to adapt to its environment. For
example, the cell does not produce enzymes needed to catabolize
lactose unless lactose is available. Also, the cell does not
produce enzymes needed for the synthesis of the amino acid
tryptophan when tryptophan is available.
Some
regions of DNA are specifically involved in regulating
transcription. These regulatory genes can control the synthesis of
specific enzymes. Sometimes gene expression is not subject to
specific genetic regulatory control. In these cases, the
enzymes coded for by such regions of the DNA are constitutive,
that is,
they are continuously synthesized. In contrast to constitutive
enzymes, some enzymes are synthesized only when the cell requires
them. Some such enzymes are inducible,
that is,
made only in response to a specific inducer substance. Others are
repressible,
that is,
made unless stopped by the presence of a specific repressor
substance.
Operons
In
1961 Francois Jacob and Jacques Monod put forth a hypothesis that
induction and repression were under the control of specific
proteins. Such proteins would be coded for by regulatory genes. They
proposed that regulatory genes were closely associated with the
structural genes that code for the enzymes in specific metabolic
pathways. Often, several enzymes that have related functions are
controlled by the same regulatory gene. Called the operon
model,
the
mechanism proposed by Jacob and Monod explains how cells are
able to coordinate the expression of genes with related functions.
An
operon
is
a cluster of adjacent genes on the chromosome that is controlled by
one promoter site. Transcription starting at that promoter site
results in the formation of an mRNA coding for several polypeptides.
Such an mRNA is said to be polycistronic,
meaning
that it codes for more than one
polypeptide.
An operator gene within the operon acts like a switch, turning on
and off the transcription of structural genes. Either all or none of
the genes of the operon are expressed. This is achieved at the level
of transcription by controlling the production of the polycistronic
mRNA. Induction and repression of genes in an operon are based on
whether or not a regulatory repressor protein binds at a
regulatory gene of the DNA, called the operator.
If the repressor protein binds to the operator, it blocks
transcription of the succeeding structural genes.
Some
operons are regulated by positive control, which involves the
binding of a regulator protein to DNA and the stimulation of gene
expression. Others are regulated by negative control, which involves
binding of a regulator protein to DNA and the shutting down of
gene expression.
Regulating
the Metabolism of Lactose—the lac
Operon
The
lac
operon coordinates the expression of three enzymes that are
specifically synthesized by Escherichia
coli for
the metabolism of lactose. These enzymes are: /3-galactosidase,
galactoside permease, and trans- acetylase. /3-galactosidase cleaves
the disaccharide lactose into the monosaccharides galactose and
glucose. Galactoside permease is required for the transport
of lactose across the bacterial plasma membrane. The role of
transacetylase is not yet established. The structural genes that
code for the production of these three enzymes occur in a contiguous
segment of DNA.
The
operon for lactose metabolism is called the lac
operon
(FIG.
7-23). The lac
operon includes a promoter region where RNA polymerase binds,
an operator region where the repressor protein attaches, and
three structural genes that code for three proteins that are
involved in lactose metabolism. In addition, there is a
regulatory gene at another location that codes for the synthesis of
a repressor protein. In the absence of lactose, this repressor
protein binds to the operator region of the DNA. The operator
region occurs between the promoter and the three structural
genes. The binding of the repressor protein at the operator region
blocks the transcription of the structural genes. This means that in
the absence of lactose, the three structural lac
genes are not transcribed.
The
operator region is adjacent to or overlaps the promoter region. The
binding of the repressor protein at the operator region
interferes with the binding of RNA polymerase at the promoter
region. The inducer binds to the repressor protein so that it
is unable to bind at the operator region. Thus in the presence
of an inducer that binds with the repressor pro-
214
FIG.
7-23 The lac
operon controls the utilization of lactose. Three structural genes
under the control of the lac
promoter (P lac)
code for the synthesis of the enzymes needed for lactose
utilization. These enzymes are made only when lactose is present.
tein,
transcription of the lac
operon is not blocked and the synthesis of the three structural
proteins needed for lactose metabolism proceeds. The lac
operon
is typical of operons that control catabolic pathways; only in the
presence of an appropriate inducer is the system turned on.
Catabolite
Repression
When
more than one carbon source such as glucose and lactose is available
at the same time, the cell will I use the simpler substance first.
Thus glucose is used before lactose. The cell turns on the genes for
glucose metabolism and does not turn on (represses) the genes for
lactose utilization. This type of repression is called catabolite
repression.
It
regulates the expression of multiple genes that are under the
control of different promoters. Only some genes are controlled I by
catabolite repression.
Catabolite
repression acts via the promoter region I of DNA. This is the region
where RNA polymerase binds to initiate transcription (FIG. 7-24). To
effi- ; dently bind to the promoter region, RNA polymerase
requires a protein called the catabolite
activator I
protein.
The catabolite activator protein, in turn, can
not
bind to the promoter region unless it is bound to cyclic adenosine
monophosphate (cAMP).
In
the absence of glucose, cAMP is synthesized from ATP by enzymatic
action. This maintains an adequate supply of cAMP to permit the
binding of RNA polymerase to the promoter region. Thus, when glucose
levels are low, cAMP stimulates the initiation of many inducible
enzymes.
In
the presence of glucose, cAMP levels are greatly reduced. Thus, when
glucose is being metabolized, there is not enough cAMP for the
catabolite activator protein to bind to promoter region.
Consequently, RNA polymerase does not bind to the promoters, and
transcription at a number of regulated structural genes ceases in a
coordinated manner. Thus, in the presence of an adequate
concentration of glucose, a number of metabolic pathways involved in
the breakdown of carbohydrates are simultaneously shut off. For
example, when glucose is available for catabolism in the
glycolytic pathway, disaccharides and polysaccharides are not
metabolized because of catabolite repression.
By
regulating the metabolism of more complex carbohydrates, the cell
conserves its metabolic resources.
REGULATION
OF GENE EXPRESSION 215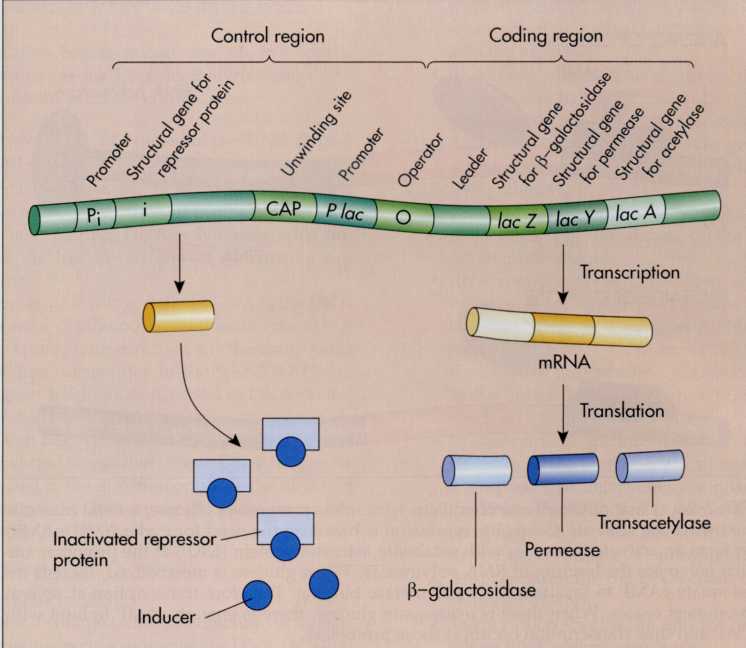
FIG.
7-24 Catabolite
repression explains why, in the presence of glucose, several
catabolic pathways are shut off. Catabolite repression is based on
the need for cyclic AMP (cAMP) to form an activated complex with
catabolite activator protein (CAP) at the promoter site that
enhances the binding of RNA polymerase. When glucose is metabolized,
there is inadequate cAMP to facilitate RNA polymerase binding.
Therefore transcription at several promoters ceases. When there is
inadequate glucose, there is enough cAMP to bind with CAP and thus
transcription occurs at those promoters.
SUMMARY
Molecular
Basis of Heredity (pp. 192-194)
Frederick
Griffith, Oswald Avery, Alfred Hershey, and Martha Chase made
important contributions to the discovery that the genetic
information of a cell is stored within its DNA macromolecules.
Structure
of DNA (pp. 194-198)
Nucleotides—Building
Blocks of the Genetic
Code
(p. 194)
DNA
is composed of nucleotides that are linked together. A
nucleotide consists of a nucleic acid base, a deoxyribose sugar,
and a phosphate group.
Four
nucleic acid bases occur in DNA: cytosine, guanine, adenine,
and thymine. The nucleotides are linked by strong covalent bonds.
Nucleotides are linked by 3'-5' phosphodiester linkages. At the
ends of the DNA strand, there are no linkages and free hydroxyl
groups are present. One end has a free hydroxyl group at the
3-carbon position of the monosaccharide (3'-OH end); the other
end of the strand has a free phosphate group at the 5-carbon
position of the monosaccharide (5'-P free end). This gives DNA
directionality.
Chains
of Nucleotides—Directionality of
DNA
(pp. 194-196)
DNA
is a double helix molecule composed of two polynucleotide chains.
The chains are held together by hydrogen bonding between
complementary nucleotide bases.
DNA
Double Helix—Complementarity (pp. 196-198)
The
complementary base pairs are adenine and thymine, which are held
together by two hydrogen bonds, and guanine and cytosine, which are
held together by three hydrogen bonds. This complementarity
establishes the basis for the double helix and the accurate
replication of DNA.
Replication
of DNA (pp. 198-202)
Replication
of the hereditary information involves synthesizing new DNA
molecules that have the same nucleotide sequences as those of the
parent organism. The two chains of the DNA double helix are
complementary and the nucleotide sequence in one chain
specifies the sequence in the other.
DNA
chains are complementary and antiparallel; one chain has the
3'-OH free end and its I complementary chain has the 5'-P free end.
Semiconservative
DNA Replication (pp. 198-199)
DNA
replication is semiconservative, that is, a parent chain remains
intact and a new complementary chain is assembled for each one.
Thus each new DNA f macromolecule is half old and half new.
Steps
in DNA Replication (pp. 199-201)
DNA
replication begins when the double helix un- winds to form a
replication fork, separating the chains to serve as templates.
The
parental DNA is pulled apart at the replication fork, providing
space for free nucleotides to align op
216
CHAPTER
7 MICROBIAL GENETICS: REPLICATION AND EXPRESSION OF GENETIC
INFORMATION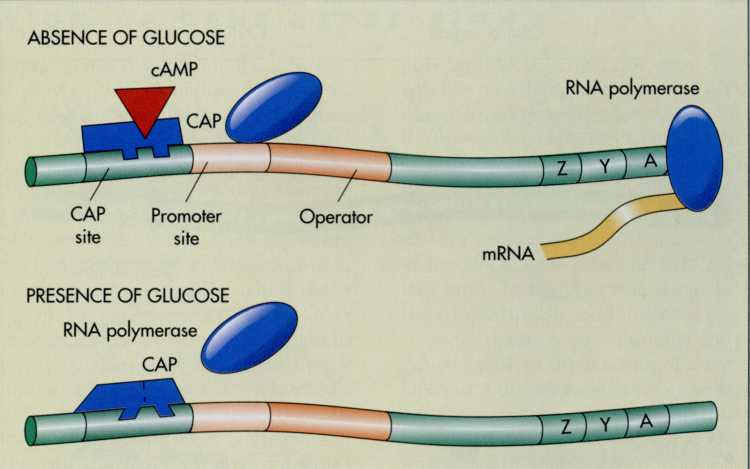
posite
their complementary bases for the synthesis of new chains.
DNA
replication begins at only one site in bacteria. DNA gyrase twists
the DNA as the replication fork moves around the bacterial
chromosome.
Formation
of a New Chain of Nucleotides—DNA Polymerase (pp. 201-202)
DNA
polymerase links the nucleotides by forming phosphodiester bonds
after the nucleotides are aligned by base pairing. DNA polymerase
adds nucleotides to the free 3'-OH end of an existing
nucleotide chain.
The
continuous, or leading, chain of DNA is the DNA chain that can be
continuously synthesized because it runs in the appropriate
direction for the continuous addition of free nucleotides to the
free 3'-OH end. The lagging, or discontinuous, strand of DNA cannot
be synthesized continuously because initiation of its replication
can begin only after the double helix has already unwound somewhat.
Short DNA fragments are synthesized in the direction opposite the
direction in which the parent DNA unwinds. These fragments are
joined together by ligases.
Mutations
(pp. 203-205)
Types
of Mutations (p. 203)
A
mutation is a change (addition, deletion, or substitution) in
the nucleotide sequences of DNA. A lethal mutation results in the
death of a microorganism or in its inability to reproduce; a
conditionally lethal mutation exerts its effect only under
certain environmental conditions; an unconditionally lethal
mutation is lethal regardless of environmental conditions.
Temperature-sensitive mutations alter the range of
temperatures over which the microorganisms may grow.
Nutritional mutations alter the nutritional requirements for
the progeny; nutritional mutants (auxotrophs) require growth
factors not needed by the parental (prototrophic) strain.
Factors
Affecting Rates of Mutation (p. 205)
Mutagens
are chemicals that increase the rate of mutation. High-energy
ionizing radiation causes mutation and can be used for
sterilizing objects. Ultraviolet light can cause mutations by
producing thymine dimers.
Expression
of Genetic Information (pp. 205-213)
The
genotype represents the total informational capacity of the
cell. It is mediated by DNA. The phenotype, the discernible
characteristics of an organism, including the functional appearance
and activities of the cell, is mediated by proteins.
The
sequence of nucleotides within the DNA determines the sequence
of amino acids in the protein molecules of the cell.
Genes
(pp. 206-207)
A
gene is a segment of the genetic material that has a specific
function. Structural genes code for the synthesis of RNA and
proteins, respectively determining the sequences of nucleotides and
amino acids in these
macromolecules.
Regulatory genes control cell activity by specifying when
particular structural genes are actually expressed.
Prokaryotic
cells have a single chromosome and therefore are haploid.
Eukaryotic cells generally have pairs of matching chromosomes,
making them diploid. In homozygous cells the genes at a locus are
identical copies; in heterozygous cells the genes differ.
RNA
Synthesis (pp. 207-210)
Protein
synthesis involves two stages: transcription to form RNA and
translation of the RNA to form a polypeptide chain.
RNA
contains ribose, phosphate, adenine, uracil, cytosine, and
guanine. There are three types of RNA. Ribosomal RNA is a
structural component of ribosomes. Messenger RNA carries the
information from the DNA to the ribosome. Transfer RNA helps align
amino acids during protein synthesis in the order specified by
mRNA.
In
transcription, the information in the DNA is transferred to
RNA. During transcription, DNA serves as a template that determines
the order of the bases in the RNA. The RNA that is formed by
transcription is complementary to the DNA. RNA polymerase links the
bases, forming 3'-5' phosphodiester bonds. The template strand is
the DNA chain that codes for the synthesis of RNA.
Transcription
begins at specific promoter regions where RNA polymerase binds.
The
sequence of nucleotides in prokaryotic mRNA corresponds exactly
with the sequence of nucleotides in DNA. Eukaryotic genes are split
genes, that is, the sequence of nucleotide bases in the mRNA is not
complementary to the specific contiguous linear sequence of
bases in the DNA. Eukaryotic RNA (heterogeneous nuclear RNA)
must be extensively modified after transcription from DNA to
form mRNA.
Protein
Synthesis—Translation of the Genetic
Code
(pp. 210-213)
In
translation, mRNA is used to establish the sequence of amino
acids that make up the protein. Translation occurs at the
ribosomes.
Translation
is a directional process. mRNA is read in a 5'-P to 3'-OH
direction. Polypeptides are synthesized from the amino
terminal to the carboxyl terminal end.
The
genetic code has 64 possible codons; each codon is a triplet
containing three nucleotides. There is more than one codon for most
amino acids, and different codons can specify the same amino acid.
Nonsense
codons are ones for which there are no amino acids; the nonsense
codons signal termination of synthesis of a polypeptide chain.
The
ribosome moves along the mRNA, exposing one codon at a time. As
each triplet is exposed by the ribosome, a transfer RNA (tRNA)
brings the specified amino acid to the ribosome; the tRNA has an
anticodon region that is complementary to the codon and is
responsible for bringing the correct amino acid specified by the
codon. The ribosome moves to the next triplet and the process is
repeated.
SUMMARY 217
218
CHAPTER
7 MICROBIAL GENETICS: REPLICATION AND EXPRESSION OF GENETIC
INFORMATION
Translocation
is the movement of mRNA, tRNA, and the polypeptide chain along the
ribosome.
Regulation
of Gene Expression (pp. 214-216)
The
expression of genetic information can be regulated at the
level of transcription.
Constitutive
enzymes are continuously synthesized at a constant rate and are not
regulated. Inducible enzymes are made only at appropriate
times, e.g., when synthesis is induced by appropriate factors.
Operons
(pp. 214-215)
The
operon model of gene control explains the basis of control of
transcription. An operon consists of structural genes that contain
the code for making proteins; an operator region, which is the
site where repressor protein binds and prevents RNA
transcription; and a promoter region, which is the site where
RNA polymerase binds. It is also controlled by a regulatory
gene, which codes for the repressor protein.
The
lac
operon regulates the utilization of lactose. In the presence of
lactose an inducer binds to a repressor protein, preventing it from
binding to the operator region of the operon; this results in
derepression of lac
operon,
and structural genes needed for the utilization of lactose are
transcribed until the lactose has been broken down.
Catabolite
Repression (pp. 215-216)
Catabolite
repression is a generalized type of repression. Catabolite
repression supersedes the control exerted by the operator
region. Catabolite repression acts via promoter region of DNA by
blocking the normal attachment of RNA polymerase; a catabolite
activator protein is needed to bind RNA polymerase to promoter
region and cAMP is required for efficient binding to occur. In the
presence of glucose, the amount of cAMP is reduced; therefore the
catabolite activator protein cannot bind to promoter, and
transcription is unable to occur.
CHAPTER
REVIEW
Review
Questions
Explain
the difference between a gene and a chromosome.
What
is the difference between genotype and phenotype?
What
is the relationship between DNA and heredity?
What
is the genetic code?
What
is a mutagen?
What
is a mutation?
How
is DNA replicated in bacterial cells?
Describe
the process of protein synthesis.
Define
induction and explain how it regulates gene expression in
bacteria.
Define
catabolite repression and explain how it regulates gene
expression in bacteria.
What
are the different types of mutations?
Describe
how the Ames test is used to detect carcinogens.
Compare
and contrast the storage of genetic information in a
prokaryotic and a eukaryotic cell.
Compare
and contrast the expression of genetic information in a
prokaryotic and a eukaryotic cell.
What
is DNA gyrase and what role does it play in DNA replication?
How
could DNA gyrase be used as a target for an antimicrobial
agent for the treatment of disease?
How
would you go about increasing the rate of mutations?
How
could you recognize the occurrence of a mutant?
How
could you design an experiment to select mu- I tants?
How
does the structure of DNA relate to the ability of this molecule to
serve as the universal hereditary molecule of all living
organisms? Why is fidelity essential for replication of DNA? How
does a bacterial cell replicate its DNA and make very few errors in
the process? What is the consequence of making an error during DNA
replication?
Why
is it so important for the bacterial cell to regulate the
expression of its genes? What are the advantages and disadvantages
of bacterial genes being organized into operons? Why are
operons for catabolic pathways normally inducible (turned on by an
inducer) and those for biosynthetic pathways normally
repressible (turned off by a repressor).
Why
can eukaryotic cells have split genes? What roles might introns
play in the eukaryotic cell?
DNA
has the sugar deoxyribose and RNA has the sugar ribose. Compared to
deoxyribose, ribose has an extra hydroxyl group. The extra hydroxyl
group tends to help break phosphate bonds. How would this affect
the relative stability of RNA and DNA in a cell? How would this
difference in stability be related to the different functions of
RNA and DNA in a cell? What are the essential functions of DNA and
RNA in a cell?
Do
all substances that cause mutations in bacteria cause cancer in
humans? How could you go about determining whether foods you
eat contain substances that might be mutagenic or carcinogenic?
CRITICAL
THINKING QUESTIONS
Readings
Alberts
B, D Bray, J Lewis, M Raff, K Roberts, JD Watson: 1989. Molecular
Biology of the Cell,
New York, Garland Press.
Comprehensive
text covering the molecular basis of heredity.
Bishop
JE and M Waddles: 1990. Genome:
the Story of the Most
As- lonishing
Scientific Adventure of our Time—the Attempt to Map all the Genes
in the Human Body,
New York, Simon and Schuster.
Discusses
the government-financed program to map every gene in human DNA, the
medical, ethical, and scientific questions this effort raises, and
searches for the genes that cause specific diseases.
Brock
TD: 1990. The
Emergence of Bacterial Genetics,
Cold Spring Harbor, New York; Cold Spring Harbor Laboratory Press.
Definitive
history of the development of bacterial genetics as we know it
today.
Cairns
], G Stent, J Watson (eds.): 1966. Phage
and the Origins of Molecular Biology,
Cold Spring Harbor, New York; Cold Spring Harbor Laboratory Press.
Collection
of essays by the founders of and converts to molecular genetics.
Gives a sense of history in the making—the emergence of insights,
the wit, the humility, the personalities of the individuals
involved.
Dahlberg
AE: 1989. The functional role of ribosomal RNA in protein
synthesis, Cell
57:525-529.
Thorough
discussion of the role of RNA in transferring information from DNA
to proteins.
Darnell
J: 1985. RNA, Scientific
American
253(4):68-78.
This
article suggests that while RNA now functions in an informational
capacity, it may have been the original genetic material.
Darnell
J, H Lodish, D Baltimore: 1990. Molecular
Cell Biology,
New York, Scientific American Books.
Well-illustrated
discussion of the molecular biology of the cell.
Dubos
R: 1976. The
Professor, the Institute, and DNA: Oswald T. Avery, his Life
and Scientific Achievements,
New York, Rockefeller University Press.
The
stories of the discoverer that hereditary characteristics are
transmitted by molecules by DNA and his institution, the
Rockefeller University.
Feldman
M and L Eisenbach: 1988. What makes a tumor cell I metastatic?
Scientific
American
259(5):60-85.
Discusses
the molecular basis for transformation to cancerous cells.
Felsenfeld
G: 1985. DNA, Scientific
American
253(4): 58-78.
It
is the double-helical structure of DNA that allows it to interact
with regulatory proteins and other molecules to transfer its
hereditary message.
Genetics:
Readings from Scientific American:
1981. San Francisco, W. H. Freeman.
Collection
of articles on molecular biology.
Grunstein
M: 1992. Histones as regulators of genes, Scientific
Amer- |
ian
267(4):68-74B.
A
discussion of the role played by histones in regulating gene
expression through repression and activation of genes.
Hawkins
JD: 1991. Gene
Structure and Expression,
Cambridge, England; Cambridge University Press.
Presents
recent ideas and techniques in molecular biology as related to
genetics so that students will be able to understand further
advances as they occur.
Innis
MA, DH Gelfand, JJ Sninsky, TJ White (eds.): 1990. PGR
Protocols: A Guide to Methods and Applications,
San Diego, Academic Press.
Authoritative
book describing ways in which PCR can be used, including medical,
environmental, and forensic applications.
Maloy
SR, JE Cronan Jr, D Freifelde: 1994, Microbial
Genetics,
ed. 2, Boston, Jones and Bartlett.
Comprehensive
text covering classical and molecular genetics of microorganisms.
McCarty
M: 1987. The
Transforming Principle: Discovering that Genes are Made of DNA,
New York, W. W. Norton.
Describes
the discovery of genetic transformation.
McKnight
SL: 1991. Molecular zippers in gene regulation, Scientific
American
264(4):54-64.
Recurring
copies of the amino acid leucine in proteins can serve as teeth that
zip protein molecules together. This zipper may play a role in
turning genes on and off.
Mullis
KB: 1990. The unusual origin of the polymerase chain reaction,
Scientific
American
262(4):56-61, 64-65.
The
description of the polymerase chain reaction and its discovery
by its discoverer and winner of the 1993 Nobel Prize in Chemistry.
Olby
R: 1974. The
Path to the Double Helix,
Seattle, University of Washington Press.
The
early history of molecular biology is reviewed.
Parker
J: 1989. Errors and alternatives in reading the universal genetic
code, Microbiological
Reviews
53(3): 273-298.
Concerns
the types of errors and alternate readings, the frequencies with
which they occur, and some of the factors that influence these
frequencies.
Prescott
D: 1988. Cells,
Boston, Jones & Bartlett.
Chapter
8 contains an excellent introduction to protein synthesis. Ptashne
M: 1989. How gene activators work, Scientific
American 260(1):
41-47.
Molecular
biologists are using what they have learned about turning bacterial
genes on and off to study genetic regulation in higher organisms.
Radman
M and R Wagner: 1988. The high fidelity of DNA duplication,
Scientific
American
259(2):40-46.
Reviews
DNA replication and the systems that ensure accuracy of information
duplication.
Venitt
S and JM Parry (eds.): 1985. Mutagenicity
Testing,
Oxford University Press.
Reviews
the use of microbial test systems identifying mutagens. Watson
J: 1978. The
Double Helix,
New York, Atheneum.
Highly
personal view of scientists and their methods, interwoven into an
exciting account of how DNA structure was discovered.
Watson
JD, N Hopkins, J Roberts, J Steitz, A Weiner: 1987. Molecular
Biology of the Gene,
Menlo Park, CA, Benjamin/Cummings.
A
comprehensive review of molecular biology.
Weintraub
HM: 1990. Antisense RNA and DNA, Scientific
American
262(l):40-46.
A
discussion of a fascinating mechanism of regulating gene
expression.
Zubay
G: 1987. Genetics.
Menlo Park, California, Benjamin/Cummings.
CHAPTER
REVIEW 219