Microbial
Genetics:
Replication
and Expression of Genetic Information
Molecular
Basis of Heredity 192 Structure of DNA 194
Nucleotides—Building
Blocks of the Genetic Code Chains of Nucleotides—Directionality of
DNA DNA Double Helix—Complementarity Historical
Perspective:
Discovering the Structure of DNA I
Replication of DNA 198
Semiconservative
DNA Replication Steps in DNA Replication Unwinding the DNA Double
Helix—Replication Forks
Methodology:
Polymerase Chain Reaction (PCR) Formation of a New Chain of
Nucleotides—DNA Polymerase I
Mutations 203
Types
of Mutations Methodology:
Ames Test Factors Affecting Rates of Mutation I
Expression of Genetic Information 205 Genes
Historical
Perspective:
One Gene—One Polypeptide RNA Synthesis Ribonucleic Acid
(RNA)—Functions and Types Transcription ; Initiation and
Termination of Transcription Synthesis of mRNA in Prokaryotic and
Eukaryotic Cells—Split Genes Protein Synthesis—Translation of
the Genetic Code mRNA and the Genetic Code tRNA and Polypeptide
Formation Forming the Polypeptide Regulation
of Gene Expression 214 Operons
Regulating
the Metabolism of Lactose—the lac
Operon
Catabolite Repression
Discover
the underlying mechanisms of heredity and the biochemical events
that enable the passage of hereditary information.
Examine
the properties of DNA (deoxyribonucleic acid), the universal master
molecule of life that stores genetic information in all
cells—bacterial, human, and other.
See
how DNA molecules are replicated so that hereditary information can
be passed from one generation to the next.
Discover
how all of the properties of an organism are determined at the
molecular level.
Learn
how genetic expression occurs, seeing how information in the DNA is
transferred through RNA (ribonucleic acid) molecules to proteins.
Learn
the following key terms and names:
Ames
test nonsense codons
anticodon nucleic
acid bases
auxotrophs nucleotides
base
pairing operon model
catabolite
repression phenotype
codon polycistronic
diploid polymerase
chain reaction
DNA
gyrase (PCR)
DNA
polymerase promoter
double
helix prototroph
exons regulatory
genes
frame-shift
mutations replica plating
genes replication
fork
genotype ribosomal
RNA (rRNA)
haploid RNA
polymerase
heterozygous semiconservative
hnRNA
(heterogeneous replication
nuclear
RNA) split genes
homozygous structural
genes
inducible template
strand
introns thymine
dimer
lethal
mutation transcription
mutagens transfer
RNA (tRNA)
mutation translation
191
Preview
to Chapter 7
Chapter
Outline

In this chapter we will:
MOLECULAR
BASIS OF HEREDITY
In
1928 a British microbiologist, Frederick Griffith, was trying to
develop a vaccine against pneumonia. He was working with two
different strains of the causative bacterium Streptococcus
pneumoniae
(FIG. 7-1). One strain was pathogenic, killing the mice injected
with it. The other strain was nonpathogenic. The two strains
differed in appearance when viewed under the microscope. The
nonpathogenic strain appeared rough and was not surrounded by a
capsule. The pathogenic strain appeared smooth, surrounded by a
polysaccharide capsule. When Griffith injected heat-killed cells of
this smooth, pathogenic strain of S. pneumoniae
into a mouse, the mouse survived because the dead bacteria were
unable to establish an
infection
in the mouse. However, when he injected a mouse with living cells of
the rough nonpathogenic strain, together with dead smooth bacteria,
knowing that neither of them could cause disease alone, the mouse
died. Unlike the live, rough bacteria he in- ; jected, the bacteria
he isolated from the dead mouse appeared smooth and surrounded by a
capsule.
This
was a most puzzling observation. Griffith reasoned that genetic
material from the heat-killed bacteria had somehow entered the
living nonpathogens [ and transformed them into pathogenic bacteria.
He postulated that heat could kill the pathogenic cells without
destroying the substance containing their f hereditary information,
which included instructions [
FIG.
7-1 The
transformation of Streptococcus
pneumoniae
shows how the properties of a bacterial strain can be altered by a
hereditary substance (later identified as DNA). When cells of S.
pneumoniae
are heat killed they leak DNA, which can be picked up by other cells
and incorporated into the genetic information of those cells. In
this manner, avirulent (nonpathogenic) strains of S.
pneumoniae
that lack the gene for capsule production (virulence factor that
contributes to their ability to cause fatal disease) can acquire the
gene (DNA) that encodes for capsule production. When this occurs, an
avirulent noncapsule-producing strain of S.
pneumoniae
is transformed into a virulent strain that produces a capsule.
192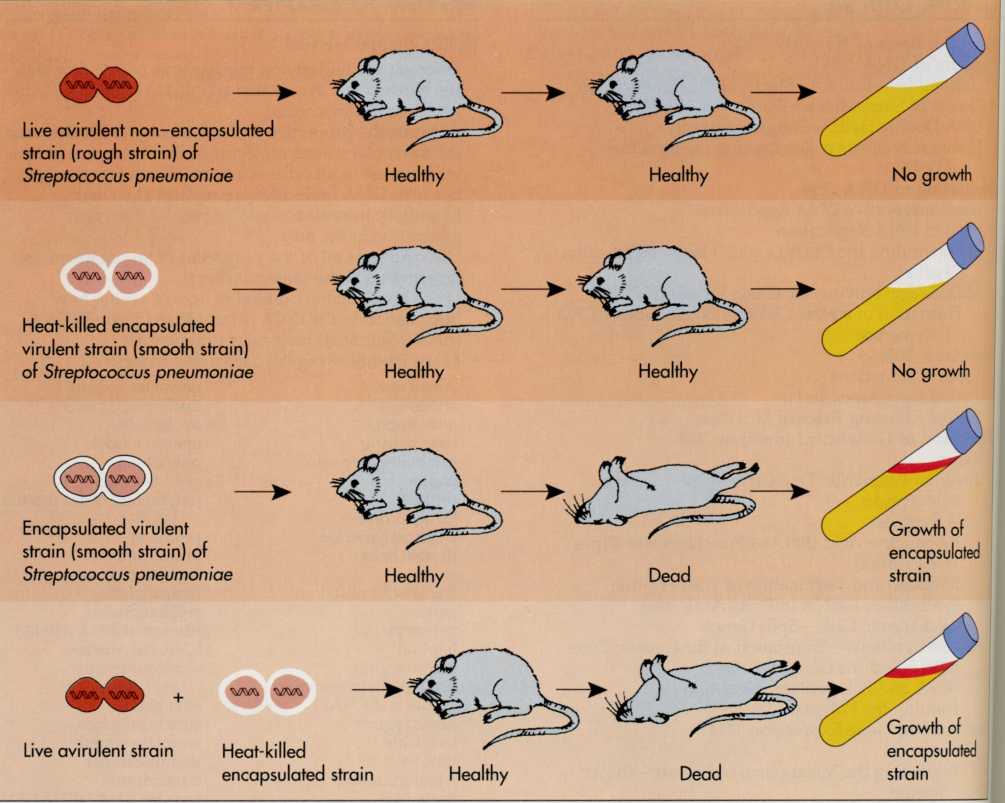
MOLECULAR
BASIS OF HEREDITY 193
on
how to cause infection and disease. Griffith had, in fact, observed
the movement of hereditary material from one cell to another.
The chemical that transmitted the hereditary information for
causing disease leaked from the dead pathogens and was picked up by
the living bacteria, transforming them into pathogens when it became
part of their hereditary material.
Other
scientists then began to investigate the specific chemical
substance that caused the transformation of a nonpathogen to a
pathogen. They were looking for the molecular basis for
heredity. Chemical analyses narrowed the possible hereditary
molecules to either proteins or nucleic acids. Most scientists
hypothesized that proteins were the basis of heredity because
their essential roles in metabolism were known. The specific
chemical nature of the transforming material observed by
Griffith, however, remained a puzzle until 1944 when Oswald
Avery and his co-workers were able to demonstrate the chemical
nature of the substance that transformed nonpathogenic S.
pneumoniae
to pathogenic S.
pneumoniae.
Avery
hypothesized that a nucleic acid, deoxyribonucleic acid (DNA),
rather than protein was the hereditary molecule. He designed
experiments to prove this. In Avery's experiments the transforming
principle of S.
pneumoniae,
which had been shown to be predominantly DNA with a trace of
protein, was treated sequentially with an enzyme that destroys
protein and an enzyme that destroys DNA (FIG. 7-2). Avery observed
that the protein-destroying enzyme did not affect the ability of the
material to transform nonpathogenic S.
pneumoniae
into pathogenic S. pneumoniae,
whereas treatment with the DNA-destroying enzyme eliminated such
transformation. Based on these observations, Avery concluded that
the transforming principle must be DNA.
Despite
this quite convincing demonstration, the scientific community was
not ready to accept that DNA was the universal hereditary molecule.
Most scientists remained convinced that proteins would eventually be
shown to be the basis of heredity for organisms other than bacteria.
Another set of experiments conducted with bacteriophage
(viruses that replicate within bacterial cells), however, added
convincing evidence that nucleic acids, not proteins, are the
source of hereditary information. These experiments, conducted
in 1952 by Alfred Hershey and Martha Chase, examined the replication
of bacteriophage T2. Although bacteriophage are not living
cells, they were known to contain DNA and protein, making them good
simple models to examine whether it is protein or DNA that carries
hereditary information.
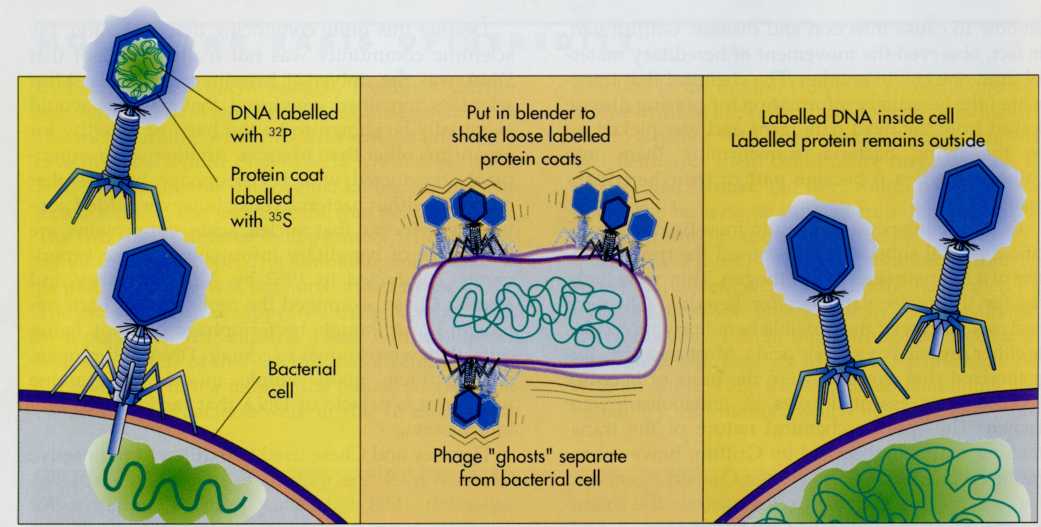
Hershey
and Chase used two different radioactive labels to track the
movement of protein and DNA separately (FIG. 7-3). Most proteins
contain sulfur but none contain phosphorus. Thus the radioactive
isotope 35S can be used to label the bacteriophage
protein. DNA contains phosphorus but no sulfur, so they used the
radioactive isotope 32P to label the bacteriophage
DNA. Thus Hershey and Chase cleverly devised a method for following
both the DNA and protein components of bacteriophage T2. When they
added bacteriophage that had been labelled with 3SS to a
culture of growing cells of the bacterium Escherichia
coli,
they observed that the 35S label remained outside of
the bacterial cells. Thus protein did not enter the bacterial cells.
In contrast, when they similarly added 32P-labelled
bacteriophages, the 32P label entered the interior of the
bacterial cells. This indicated that DNA was the material that
entered the cells and therefore must be the substance that carried
the hereditary information. The progeny bacteriophages produced
from the replication of the original
FIG.
7-2 To
prove that the hereditary substance was DNA, enzymes that degrade
proteins were added to cell extracts. These enzymes did not
eliminate transformation, showing that the substance was not a
protein. In contrast, the addition of a DNA-destroying enzyme
eliminated transformation.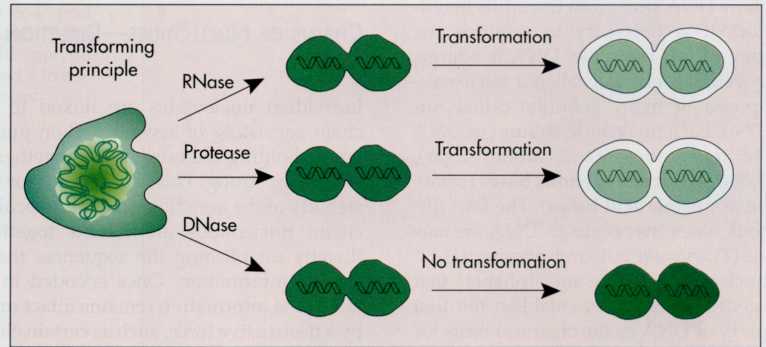
FIG.
7-3 Hershey
and Chase demonstrated that nucleic acids are the hereditary
substances of viruses. In their experiments 32P was
used to label nucleic acids and 35S was used to label
proteins. The 35S remained outside of the host cell,
whereas the 32P entered the cell. This indicated that the
32P-labelled nucleic acid carried the hereditary
information.
bacteriophage
contained 32P and not 35S, indicating further
that the hereditary material passed from one generation to the next,
was, in fact, DNA. Although subsequent experiments have shown that
another nucleic acid (ribonucleic acid [RNA]) sometimes is the
hereditary substance for viruses, it was now
STRUCTURE
OF DNA
Nucleotides—Building
Blocks of the Genetic Code
To
understand how DNA stores and transmits hereditary information,
it is necessary to examine the chemical structure of this molecule.
DNA is a large, high-molecular-weight molecule, called a
macromolecule.
It is composed of many subunits called nucleotides
(FIG.
7-4). Each nucleotide subunit of DNA has three parts: deoxyribose (a
5-carbon sugar), phosphate, and one of four nitrogenous
bases
(sometimes
referred to as nucleic
acid bases).
The four
different nitrogenous bases that occur in DNA are adenine
(A), thymine
(T), guanine
(G), and cytosine
(C).
These
four nucleotides are like an "alphabet" that makes up the
genetic code. They establish the first important property of DNA as
the chemical basis for heredity—the ability to encode the genetic
information. This is achieved by linking the nucleotides in a
specific order—much as the letters of the alphabet are joined to
form words.
The
hereditary information is coded by the order in which the four
different nucleotides occur within the DNA macromolecule.
Chains
of Nucleotides—Directionality of DNA
Individual
nucleotides are linked to form a long chain consisting of several
million nucleotides. The bonds holding the nucleotides together are
covalent and hence strong. This is important for the long-term
stability of the hereditary macromolecule. Within this chain,
nucleotides are locked together in order, thereby establishing the
sequences that encode the genetic information. Once encoded in the
chain of DNA, the information remains intact unless acted on by a
destructive force, such as certain chemicals or radiation.
The
chemical bonds holding the chains of nucleotides together are
called 3'-5'
phosphodiester bonds
(FIG.
7-5). They are so-named because phos-
firmly
established that DNA is the hereditary molecule for many
viruses and all living cells.
DNA
is the substance that transmits the hereditary information of many
viruses and all cellular organisms.
CHAPTER
7 MICROBIAL GENETICS: REPLICATION AND EXPRESSION OF GENETIC
INFORMATION
194