

2.2.1. Галактозилцераміди.
Галактозилцераміди (цереброзиди) – головні сфінголіпіди мозку і інших нервових тканин, але в невеликих кількостях зустрічаються і в багатьох інших тканинах. До складу галактозилцерамідів входить гексоза (звичайно це D-галактоза), яка зв'язана ефірним зв'язком з гідроксильною групою аміноспирту сфінгозину. Крім того, у складі галактозилцераміду є жирна кислота. Найчастіше це лігноцеринова, нервоновая або церебронова кислота, тобто жирні кислоти, що мають 24 вуглецевих атома.
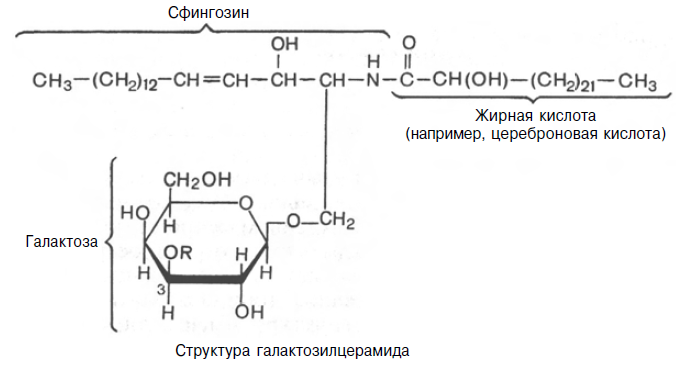
Існують сульфогалактозилцераміди, які відрізняються від галактозилцерамідів наявністю залишку сірчаної кислоти, приєднаного до третього вуглецевого атома гексози. У мозку ссавців сульфогалактозилцераміди в основному знаходяться в білій речовині, при цьому вміст їх в мозку набагато нижчий, ніж галактозилцерамідів.
2.2.1. Глюкозилцераміди.
Глюкозилцераміди – прості глікосфінголіпіди, представлені в тканинах, відмінних від нервової, причому головним чином глюкозилцерамідами. У невеликих кількостях вони є в тканині мозку. На відміну від галактозилцерамідів у них замість залишку галактози є залишок глюкози.
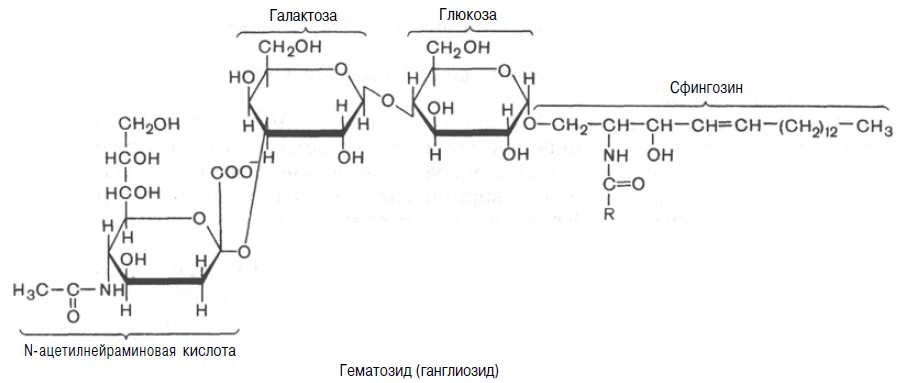
Складнішими глікосфінголіпідами є гангліозиди, що утворюються з глікозилцерамідів. Гангліозіди додатково містять одну або декілька молекул сіалової кислоти. У тканинах людини домінуючою сіаловой кислотою є нейрамінова. Крім того, замість залишку глюкози вони частіше містять складний олігосахарид.
Гангліозіди у великих кількостях знаходяться в нервовій тканині. Вони виконують рецепторні і інші функції.
Одним з простих гангліозидів є гематозид, виділений із строми еритроцитів. Він містить церамід (ацилсфінгозин), одну молекулу глюкози, одну молекулу N-ацетилнейрамінової кислоти.
2.3. Діольні ліпіди.
У природних ліпідах складноефірні зв'язки із залишками вищих жирних кислот і фосфорною кислотою, так само як і прості ефірні зв'язки із залишками вищих спиртів і вуглеводів, можуть утворювати двоатомні спирти – етандіол, а також пропандіоли, бутандіол і пентандіоли. Такі ліпіди називають діольними ліпідами.
Вони широко поширені серед рослин, тварин і мікроорганізмів як мінорні компоненти ліпідної фракції, хоча у морських безхребетних і риб доволі часто складають головну частину запасних ліпідів.
Вони також є структурними елементами мембран ендоплазматичної мережі, але не мітохондрій. Відомо 9 груп диольних ліпідів, виділених з різних об'єктів.
2.4. Орнітіноліпіди.
Орнітіноліпіди характерні для мікроорганізмів. Їх неодмінними складовими частинами є вищі жирні β–оксикислоти і амінокислота орнітин (або лізин); залежно від типу будови можуть бути присутніми також залишки етилгліколю або 1,3-пропандіолу.
Найбільш поширені в природі орнітиноліпіди побудовані таким чином:
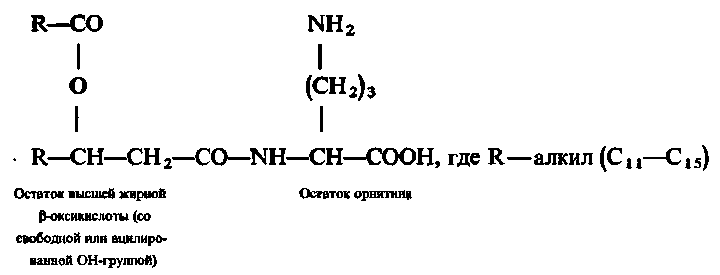
Встановлено, що орнітиноліпіди в мікробній клітині здатні замінювати фосфатидилетаноламін в якості структурного елементу цитоплазматичних мембран. Це особливо яскраво виявляється в умовах фосфатного голодування.
3. Убіхінони.
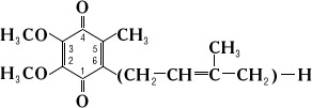
Убіхінони (від лат. ubique — скрізь, всюди і хінон), бензохінони, коферменти Q, KoQ, похідні 5-метил-2,3-диметоксіхінону, в яких до 6-го атому вуглецю приєднаний поліізопреновий ланцюг; розрізняються числом (від 6 до 10) ізопренових ланок (C5H8). Беруть участь у реакціях, що забезпечують живі клітини енергією.
Позначаються: УХn чи KoQn; у першому випадку n — число вуглецевих атомів, у другому — число ізопренових ланок (тобто, наприклад УХ30 відповідає Ko6).
Безбарвні кристалічні речовини, з максимумом поглинання при 270 мкм, нерозчинні у воді, розчинні в органічних розчинниках.
Присутні у всіх організмах: у ссавців переважає УХ50, у грибів — УХ30-50, у бактерій — УХ40, у найпростіших — УХ40-50, у комах — УХ45-50. У невеликих кількостях УХ40-50 зустрічаються в мітохондріях рослин; у хлоропластах рослин містяться близькі до убіхінонів пластохінони, у деяких бактерій – різні нафтохінони.
Біологічна дія убіхінонів основана на їх здатності до обернених окислювально-відновних перетворень. Убіхінони локалізовані в цитоплазматичних мембранах (у бактерій) та внутрішніх мембранах мітохондрій і беруть участь у перенесенні електронів і водню по дихальному ланцюгу на ділянці між флавопротеїдом і цитохромом b (див. окислювальне фосфорилювання).