

1. Виды торможения в коре головного мозга.
Процесс уменьшения или исчезновения условного рефлекса - это его торможение.
Существует 2 вида торможения:
безусловное торможение - возникает по принципу безусловного рефлекса. Особенности: безусловное торможение это врожденная форма торможения, она присуща всем особям данного вида; не нужно время для его возникновения; он может развиваться в любом отделе центральной нервной системы.
Безусловное торможение может быть:
внешнее торможение: одновременное действие света и другого сильного раздражителя приводит к торможению выделения слюны. Механизм: дополнительный внешний раздражитель вызывает новый очаг возбуждения в коре головного мозга, который является доминантным. Значение: переключение внимания с одного раздражителя на другой;
запредельное торможение: условный рефлекс подчиняется закону силы раздражения (при увеличении силы раздражителя до определенного предела идет увеличение ответной реакции). При дальнейшем увеличении силы раздражителя условные рефлексы тормозятся. Механизм: условный рефлекс резко увеличивает силу и превышает порог работоспособности нейронов коры головного мозга. Как следствие в мозговом отделе анализатора возникает запредельное торможение. Значение: предохраняет нейроны коры головного мозга от истощения.
Условное торможение - осуществляется по принципу условного рефлекса.
Особенности:
это индивидуальная, приобретенная в течении жизни реакция организма;
требует определенных условий, для осуществления его надо вырабатывать;
развивается в нейронах коры головного мозга.
Условное торможение возникает при неподкреплении условного сигнала. В коре головного мозга перестает осуществляться временная рефлекторная связь.
Причины прекращения временной рефлекторной связи:
теория Бериташвили: при неподкреплении условного сигнала действием безусловного раздражителя очаг возбуждения в корковом представительстве центра безусловного рефлекса теряет свое доминантное значение. В результате, преобладающем является возбуждение в мозговом отделе анализатора. В данном случае временная рефлекторная связь замыкается в направлении к мозговому отделу анализатора;
теория Анохина: условное торможение возникает, если в нейронах акцептора результата действия происходит несоответствие полученного результата, эталону. Формируется новый рефлекс, а старый тормозится.
Условное торможение бывает 4-х видов:
касательное - возникает при неподкреплении условного сигнала действием безусловного раздражителя. В этом случае очаг возбуждения в корковом представительстве безусловного рефлекса теряет доминирующее значение. Значение: организм избавляется от <ненужных> рефлексов;
дифференциальное значение - точное различение близких раздражителей. Механизм: в нейронах мозгового анализатора происходит дифференцировка раздражителей;
запаздывающее - в коре головного мозга возникает процесс торможения вместе возбуждения на 1-2 с, а затем вырабатывается новый условный рефлекс - изменяется временное соотношение. Значение: обеспечивает временные условные рефлексы. Этот вид торможения используется как критерий уравновешенности для определения процессов возбуждения и торможения в центральной нервной системе;
условный тормоз - условный сигнал сочетается с действием дополнительного раздражителя.
Свет + пища - через 1-2с происходит выделение слюны.
Свет + слабый звонок / нет пищи - выделение слюны.
Звонок стал тормозом. Но(!) этот звонок тормозит любой условный рефлекс данного индивида. Предполагают, что условный тормоз возникает вследствие того, что возникает дополнительный очаг, тормозящий различные условные рефлексы. Значение: основа дисциплины.
Значение торможения условного рефлекса:
отношение организма с окружающей средой становится более совершенными;
осуществляется более детальный анализ и синтез информации.
Торможение - активный, неразрывно связанный с возбуждением процесс, приводящий к задержке деятельности нервных центров или рабочих органов. В первом случае торможение называется центральным, во втором -периферическим.
В зависимости от природы физиологического механизма, лежащего в основе тормозного эффекта на условнорефлекторную деятельность организма, различают безусловное (внешнее и запредельное) и условное (внутреннее) торможение условных рефлексов.
Торможение безусловное - разновидность коркового торможения. В отличие от условного торможения наступает без предварительной выработки. Включает в себя: 1) индукционное (внешнее) торможение; 2) запредельное (охранительное) торможение.
Внешнее торможение условного рефлекса возникает под действием другого постороннего условного или безусловного раздражителя. Когда под влиянием какого-нибудь изменения внешней или внутренней среды в коре больших полушарий возникает достаточно сильный очаг возбуждения, то вследствие отрицательной индукции, возбудимость других ее пунктов оказывается пониженной - в той или иной степени в них развивается тормозное состояние.
Индукционное (внешнее) торможение - экстренное прекращение условнорефлекторной деятельности под воздействием посторонних стимулов, биологическое значение его - преимущественное обеспечение ориентировочной реакции на неожиданно возникший раздражитель. Примером такого торможения может служить следующий опыт.
У собаки выработан прочный условный рефлекс на свет электрической лампочки. Величина реакции - 10 капель слюны за 30 секунд изолированного действия раздражителя. Включение одновременно с зажиганием лампочки нового раздражителя (звонка) привело к уменьшению условного рефлекса до 1-2 капель. На повторное действие света (без включения звонка) выделилось 7 капель слюны. Испробованный еще через несколько минут условный рефлекс на свет лампочки полностью восстановился. Таким образом, под влиянием нового постороннего раздражителя произошло торможение услов¬ного рефлекса, продолжавшееся в течение некоторого времени. Источником индукционного торможения могут оказаться и раздражения, возникающие в самом организме, например сдвиги в работе пищеварительного аппарата.
При повторном действии одного и того же постороннего раздражителя вызываемый им очаг возбуждения постепенно слабеет, явление индукции исчезает и в результате прекращается тормозящее влияние на условные рефлексы.
Если изолированное действие условного пищевого раздражителя, обычно подкрепляемого через 20 секунд, продолжать 2-3 минуты, выделение слюны прекратится. То же произойдет при чрезмерном усилении раздражения. Секреция прекращается в результате развившегося торможения. Это можно доказать, пробуя также другие условные раздражители. Примененные сразу же после удлиненного во времени или чрезмерно сильного раздражения, они вызывают слабую рефлекторную реакцию вследствие иррадиации тормозного процесса на другие клетки коры.
Торможение, развивающееся в корковой клетке под влиянием длительного или сверхсильного раздражения, Павлов назвал запредельным.
Запредельное (охранительное) торможение - торможение, возникаю¬щее при действии стимулов, возбуждающих соответствующие корковые структуры выше присущего им предела работоспособности, и обеспечивающее тем самым реальную возможность ее сохранения или восстановления.
Как индукционное, так и запредельное торможение свойственно не только коре больших полушарий, но и всем другим отделам нервной системы. Существует, однако, вид торможения, встречающийся только в высшем отделе головного мозга. Такое специфическое корковое торможение Павлов назвал условным или внутренним.
Условное (внутреннее) торможение условного рефлекса носит условный характер и требует специальной выработки. Биологический смысл его в том, что изменившиеся условия внешней среды требуют соответствую¬щего адаптивного приспособительного изменения в условнорефлекторном поведении.
При выработке обычного условного рефлекса устанавливается связь раздражаемого пункта с другим возбужденным пунктом коры. При выработке условного торможения действие раздражителя связывается с тормозным состоянием корковых клеток. Один и тот же раздражитель в зависимости от того, с каким состоянием коры связывается его действие, может привести к образованию либо условного рефлекса, либо условного торможения. В первом случае он станет положительным условным раздражителем, а во втором -отрицательным.
Развитие торможения легко обнаружить в эксперименте. Так, у собаки предварительно были выработаны прочный условный рефлекс на удары метронома с подкреплением через 3 минуты и условные рефлексы на другие раздражители с подкреплением через 30 секунд. Затем метроном пускался в ход на 1 минуту и тотчас заменялся другим раздражителем, подкрепляемым через 30 секунд. При такой постановке опыта эффект действия второго раздражителя оказывался резко сниженным, то есть заторможенным. Очевидно, торможение, развившееся под влиянием, одноминутного действия метронома, захватило и другие участки коры.
Различают четыре вида внутреннего торможения: угасание, дифференцировка, условный стимул, запаздывание.
Если условный раздражитель предъявляется без подкрепления безусловным, то через некоторое время после изолированного применения условного стимула реакция на него угасает. Такое торможение условного рефлекса называется угасательным (угасание). Угасание условного рефлекса - это временное торможение, угнетение рефлекторной реакции. Спустя некоторое время новое предъявление условного стимула без подкрепления его безусловным вначале вновь приводит к проявлению условнорефлекторной реакции.
Если у животного или человека с выработанным условным рефлексом на определенную частоту звукового стимула (например, звука метронома с частотой 50 в секунду) близкие по смыслу раздражители (звук метронома с частотой 45 или 55 в секунду) не подкреплять безусловным стимулом, то условно рефлекторная реакция на последние угнетается, подавляется. Такой вид внутреннего (условного) торможения называется дифференцировочным торможением (дифференцировка). Дифференцировочное торможение лежит в основе многих форм обучения, связанных с выработкой тонких навыков.
Если условный стимул, на который образован условный рефлекс, применяется в комбинации с некоторым другим стимулом и их комбинация не подкрепляется безусловным стимулом, наступает торможение условного рефлекса, вызываемого этим стимулом. Этот вид условного торможения называется условным тормозом.
Запаздывательное торможение - торможение, наступающее тогда, когда подкрепление условного сигнала безусловным раздражителем осуществляется с большим опозданием (2-3 мин.) по отношению к моменту предъявления условного раздражителя.
Сигнальные системы
Особенности ВНД человека. Учение И.П.Павлова о 1 и 2 сигнальных системах действительности. Развитие 2 сигнальной системы у человека.
Учение И. П. Павлова о I и II сигнальных системах является логическим развитием его учения об условных рефлексах. И. П. Павлов показал, что в основе ВНД у высших животных и у человека лежат общие механизмы, но при этом существуют качественные различия. ВНД человека сформировалась исторически в процессе трудовой деятельности и необходимости общения. Проявлением ВНД человека являются речь, мышление и сознание.
I сигнальная система – это условно-рефлекторная деятельность коры больших полушарий, связанная с восприятием через рецепторы непосредственных конкретных раздражителей (сигналов) внешнего мира (свет, цвет, звук, t°…). И. П. Павлов писал: Это 1-ая сигнальная система действительности, общая у нас с животными».
II сигнальная система – это условно-рефлекторная деятельность коры больших полушарий, связанная с восприятием сигналов любого свойства (речь, жесты), причём каждый из этих сигналов имеет соответствие в I сигнальной системе и способен замкнуть рефлекс. По И. П. Павлову, чрезвычайной прибавкой к механизмам нервной деятельности является II сигнальная система, которая возникла в результате трудовой деятельности человека и появления речи. Деятельность II сигнальной системы проявляется в речевых условных рефлексах. Слово слышимое, произносимое (речь), видимое (письмо, азбука глухонемых), осязаемое (азбука слепых) является условным раздражителем, сигналом о конкретных раздражителях среды, т. е.«сигналом сигналов». «Слово, - пишет И. П. Павлов, - составило 2-ую, специальную нашу сигнальную систему действительности, будучи сигналом 1-ых сигналов». В образовании рефлексов II сигнальной системы участвуют лобные доли и мозговые центры речи. Со II сигнальной системой связана специальная человеческая особенность ВНД – способность отвлечения и обобщения сигналов, поступающих через 1-ую сигнальную систему. Сигнальное значение слова связано не с простым звукосочетанием, а с его смысловым содержанием. Для человека слово – это не только сочетание букв, но, прежде всего, форма отображения материальных явлений и предметов окружающего мира в понятиях и мыслях. II сигнальная система обеспечивает абстрактное мышление в виде умозаключений, понятий, суждений. При обобщении предметов и явлений человек открывает связи между ними (законы). II сигнальная система является высшим регулятором различных форм поведения человека в окружающей его природе и социальной среде.
С помощью II сигнальной системы, человек может получать знания и накапливать опыт без непосредственного контакта с окружающей его действительностью.
Особенности II сигнальной системы.
1) Имеется только у человека.
2) Образование условных рефлексов на базе I сигнальной системы на основе речевой деятельности.
3) Обеспечивает восприятие информации в виде символов (слов, знаков, формул, жестов).
4) В образовании речевых рефлексов участвуют лобные доли.
5) Обеспечивает абстрактное мышление человека.
Вторая сигнальная система — специальный тип высшей нервной деятельности человека, система «сигналов сигналов», идущих от общей (но не одинаковой) с животными первой сигнальной системы — ощущений, представлений, относящихся к окружающему миру. Речь, как вторая сигнальная система, как семиотическая система значимостей(см. Семиотика) — это «идущие в кору от речевых органов есть вторые сигналы, сигналы сигналов. Они представляют собой отвлечение от действительности и допускают обобщение, что и составляет наше личное, специально человеческое, высшее мышление, создающее сперва общечеловеческий эмпиризм, а, наконец, и науку — орудие высшей ориентировки человека в окружающем мире и в самом себе». И. П. Павлов (1932).
Мозг животного отвечает лишь на непосредственные зрительные, звуковые и другие раздражения или их следы; возникающие ощущения составляют первую сигнальную систему действительности.[1]
В процессе эволюции животного мира на этапе становления и начального развития вида Homo sapiens произошло качественное видоизменение системы сигнализации, обеспечивающее активное и коллективное адаптивное приспособительное поведение, создавшее многообразные, принятые в группе системы сигнализации и языки: слово, по выражению И. П. Павлова, становится «сигналом сигналов». (см.подробнее: Знаковая система). Появление второй сигнальной системы — возникновение речи и языков, сигнальных систем человека с сородичами, где условные (произвольные) сигналы индивида приобретают определенные, принятые группой значения и значимости, преобразуются в знаки языка в прямом смысле этого слова — это один из важнейших результатов многомиллионнолетней эволюции социальной жизни рода Homo, передающиеся через речевую деятельность из поколения в поколение. Биопсихологические и социальные условия формирования структур мозга (неокортекса) и образования языков подвергнуты лишь в последние полтораста лет глубокому анализу палеопсихологами Б. Ф. Поршневым, (см. его О начале человеческой истории) и антропологами. А лингвистами — лишь с открытием европейской наукойсанскрита и с появлением сравнительного языкознания индоевропейских языков (см. В.фон Гумбольдт, Фердинанд де Соссюр) .
В своей работе «Рефлекс свободы» И. П. Павлов разделяет функции сигнальных систем следующим образом:
Всю совокупность высшей нервной деятельности я представляю себе так. У высших животных, до человека включительно, первая инстанция для сложных соотношений организма с окружающей средой есть ближайшая к полушариям подкорка с её сложнейшими безусловными рефлексами (наша терминология), инстинктами, влечениями, аффектами, эмоциями (разнообразная, обычная терминология). Вызываются эти рефлексы относительно немногими безусловными внешними агентами. Отсюда — ограниченная ориентировка в окружающей среде и вместе с тем слабое приспособление.
Вторая инстанция — большие полушария… Тут возникает при помощи условной связи (ассоциации) новый принцип деятельности: сигнализация немногих, безусловных внешних агентов бесчисленной массой других агентов, постоянно вместе с тем анализируемых и синтезируемых, дающих возможность очень большой ориентировки в той же среде и тем же гораздо большего приспособления. Это составляет единственную сигнализационную систему в животном организме и первую в человеке. Анализ и синтез, осуществляемый корой больших полушарий головного мозга, в связи с наличием В. с. с. касается уже не только отдельных конкретных раздражителей, но и их обобщений, представленных в словах. В. с. с. возникла в процессе эволюции, в процессе общественного труда. Способность к обобщённому отражению явлений и предметов обеспечила человеку неограниченную возможность ориентации в окружающем мире и позволила ему создать науку. П. с. с. и В. с. с. — различные уровни единой высшей нервной деятельности, но В. с. с. играет ведущую роль. Формирование В. с. с. происходит только под влиянием общения человека с другими людьми, то есть определяется не только биологическими, но и социальными факторами. Характер взаимодействия П. с. с. и В. с. с. может варьировать в зависимости от условий воспитания (социальный фактор) и особенностей нервной системы (биологический фактор). Одни люди отличаются относительной слабостью П. с. с. — их непосредственные ощущения бледны и слабы (мыслительный тип), другие, наоборот, воспринимают сигналы П. с. с. ярко и сильно (художественный тип). Для полноценного развития личности необходимо своевременное и правильное развитие обеих сигнальных систем.
В человеке прибавляется… другая система сигнализации, сигнализация первой системы — речью, её базисом или базальным компонентом — кинестетическими раздражениями речевых органов. Этим вводится новый принцип нервной деятельности — отвлечение и вместе обобщение бесчисленных сигналов предшествующей системы, в свою очередь опять же с анализированием и синтезированием этих первых обобщенных сигналов — принцип, обусловливающий безграничную ориентировку в окружающем мире и создающий высшее приспособление человека — науку, как в виде общечеловеческого эмпиризма, так и в её специализированной форме.[2]
— Павлов И.П. "Рефлекс свободы"
В изучении В. с. с. вначале преобладало накопление фактов, характеризующих значение обобщающей функции словесных сигналов, а затем — вскрытие нервных механизмов действия слова. Установлено, что процесс обобщения словом развивается как результат выработки системы условных связей (см. Условные рефлексы); при этом имеет значение не только количество связей, но и их характер: связи, выработанные во время деятельности ребёнка, облегчают процесс обобщения. При воздействии словесных сигналов наблюдаются стойкие изменения возбудимости, большая сила, частота и длительность электрических разрядов в нервных клетках определённых пунктов коры мозга. Развитие В. с. с. — результат деятельности всей коры больших полушарий; связать этот процесс с функцией какого-то ограниченного отдела мозга невозможно. В исследованиях В.с.с. в лаборатории высшей нейродинамики и психологии высших когнитивных процессов Е. И. Бойко [1]
показана плодотворность учения И. П. Павлова о динамических временных связях В.с.с.[3] В развитие идей И. П. Павлова и Е. А. Бойко в школе Е. А. Бойко разработана общая когнитивистская модель целостного рече-мысле-языкового процесса, найдены решения сложнейших теоретических проблем психологии в ее взаимосвязях с лингвистикой, такие как вопросы соотношения языка и речи в процессах речепроизводства и речепонимания; характер связей речи с мыслью, речи с личностью говорящего; особенности развития детской речи и др. Здесь разработаны новые методы анализа публичных выступлений (интент-анализ), позволяющий в известной мере реконструировать «картину мира» говорящего — его целевые и предметные направленности, их динамику, особенности в конфликтной ситуации, в свободных условиях общения, в публичных выступлениях и др.
Существенным резервом для дальнейших исследований остаются проблемы типологии колоссальных индивидуальных различий во взаимосвязях общего и специального типов ВНД, неокортекса и эмоционально-волевой и непроизвольной регуляции деятельности и общения, пока что слабо представленных как в физиологии ВНД
Наиболее важные результаты в исследовании меж-полушарной церебральной асимметрии были получены в 60-х гг. 20 столетия амер. нейрофизиологом Р. Сперри (лауреат Нобелевской премии 1981) и его коллегами из Калифорнийского технологического института. Их опыты над пациентами с разделенным мозгом, получившие убедительную интерпретацию на основе модели переработки информации, показали, что левое полушарие полностью сохраняет способность к письму и речевому общению, к грамматически правильным ответам оно свободно оперирует символьными репрезентациями — знаками, цифрами, а также математическими формулами и др. формальными правилами. Оно способно выявлять повторяющиеся корреляции (в том числе и музыкальный ритм), но в то же время испытывает серьезные затруднения при выполнении задач на распознавание сложных образов, не поддающихся разложению на простые элементы (напр., идентификация изображений человеческих лиц). Характерно также, что у пациентов с разделенным мозгом правая рука, функционально подчиненная левому полушарию, утрачивает способность к рисованию (но не к письму), к копированию геометрических фигур, к составлению из кубиков простых композиций. Однако с этими тестами на пространственно-образное мышление гораздо успешнее справляется (особенно при выполнении двигательных задач) левая рука, функционально подчиненная правому полушарию. Это полушарие понимает элементарную речь, простые грамматические конструкции оно способно к очень ограниченной речепродукции и в состоянии справиться с элементарными аналитическими задачами.
Латерализация (разделение) и перекрещивание функций двух полушарий наблюдались исследователями также и применительно к процессам обработки зрительной и слуховой информации. Как оказалось, поле нашего зрения резко разграничено по вертикали, хотя эту границу мы субъективно не воспринимаем. Информация от правого поля зрения обоих глаз поступает в левое полушарие нашего мозга, а информация от левого поля зрения — в правое полушарие. Хотя информация, создаваемая на основе сигналов, поступающих из левого и правого полей зрения, одна и та же, обрабатывается она правым и левым полушариями по-разному. Аналогичным образом звуковые сигналы, воспринимаемые правым ухом, передаются, главным образом, в левое полушарие, и наоборот. Однако применительно к более примитивному органу чувств — обонянию — вообще не было обнаружено перекрещивания функций.
Дальнейшие исследования здоровых людей в целом подтвердили наличие Ф. а. м. и двух когнитивных типов мышления — мышления правополушарного (пространственно-образного) и левополушарного (знаково-символического). С помощью метода электроэнцефалограммы было установлено, что при выполнении тестов, требующих аналитических мыслительных стратегий (напр., устный счет), происходит активация левого полушария, в то время как правое полушарие дает на электроэнцефалограмме альфа-ритм, характерный для бездействующего полушария. Убедительные данные о наличии Ф. а. м. и двух когнитивных типов мышления были получены также с помощью метода позитронно-эмиссионной томографии. Этот метод позволил выявить и наглядно представить с помощью сложной компьютерной техники локальные зоны активности мозга, обеспечивающие переработку различных видов когнитивной информации. Так, напр., у тех испытуемых, которые пытались вспомнить какую-то музыкальную мелодию, томограф зафиксировал активность соответствующих зон правого полушария; если же при этом пользовались нотами, то наблюдалась активность левой гемисферы. Это позволило предположить, что правое полушарие неповрежденного мозга оперирует исключительно перцептивными образами и представлениями, обеспечивая ориентацию в пространстве, а левое полушарие обрабатывает информацию, представленную только в символьной, в том числе и в словесно-знаковой, форме. Однако, как показали дальнейшие эксперименты, функциональные различия между полушариями не определяются специфическими форматами внутренних репрезентаций (т.е. тем, представлена ли информация в символьной, словесно-знаковой или перцептивно-образной формах). Хотя правое полушарие и не способно к развитой речепродукции, оно все же воспринимает элементарную речь и простые грамматические конструкции, а левое полушарие может оперировать несложными перцептивными представлениями и геометрическими фигурами. Поэтому исследователи пришли к выводу, что различия между функциями полушарий и, соответственно, когнитивными типами мышления касаются, главным образом, способов переработки информации, принципов организации контекстуальной связи стимулов, доминирующих мыслительных стратегий.
Как оказалось, для пространственно-образного мышления характерны холистическая стратегия обработки многих параметров поступающей информации: оно как бы работает параллельно с несколькими входами аналогично нейрокомпьютеру. В результате происходит одновременное выявление соответствующих контекстуальных связей между различными смыслами образа или между целостными образами, «гештальтами», и создание на этой основе многозначного контекста (напр., мозаичной или калейдоскопической картины) с множественными «размытыми» связями. Конечно, весь спектр смыслов такого контекста не может быть передан с помощью вербальной системы коммуникации. Со своей стороны, знаково-символическое (логико-вербальное) мышление использует аналитическую стратегию, обеспечивая выявление только некоторых, существенных для анализа признаков и отношений, жестких причинно-следственных связей. Оно последовательно перерабатывает когнитивную информацию (вербальную и невербальную) по мере ее поступления, организуя однозначный контекст, необходимый для успешной вербальной коммуникации. Однако дальнейшие опыты показали, что при предъявлении испытуемым простейших стимулов различия между когнитивными типами мышления, касающихся стратегий обработки информации, почти полностью нивелируются. Оказалось, что когда, напр., правому и левому полям зрения предъявляли набор букв или простых геометрических фигур, знаково-символическое (логико-вербальное) мышление также обнаруживает способность к одновременной обработке информации о нескольких объектах, а пространственно-образное мышление — лишь некоторые, хотя и довольно примитивные, способности к анализу.
Были выявлены также весьма важные особенности переработки когнитивной информации, связанные с межполушарнои специализацией, с доминантностью одного из полушарий мозга. Оказалось, что наше правое полушарие, скорее всего, либо вообще не имеет прямого доступа к нашему сознательному опыту, либо этот доступ для него крайне ограничен. Есть основания полагать, что в случае доминантного левого полушария оно функционирует в качестве своего рода медиатора и интерпретатора сигналов, поступающих из правого полушария, которое не обладает развитыми вербальными способностями. Левое полушарие пытается адаптировать эти сигналы к системе личностных ориентации, установок и т.п., а иногда даже стремится блокировать их и подавлять. Это полушарие также способно порождать свою собственную модель перцептивного воображаемого пространства и своего рода «дополнительное» пространственно-образное мышление, где используются аналитические стратегии, адаптированные к нуждам манипулирования перцептивными мысленными репрезентациями — символьными изображениями, схемами, графиками и т.д., которые поддаются аналитическому разложению на более простые элементы.
Как свидетельствуют исследования анатомического строения мозга высших приматов, асимметрия височных долей левого и правого полушарий (кроме, естественно, человека) присуща только шимпанзе — определенная часть левой доли у них более развита. У человеческого эмбриона эта асимметрия возникает на двадцать девятой неделе беременности, что свидетельствует о сильной генетической предрасположенности к локализации центра управления речью именно в височной доле левого полушария. Однако у новорожденных при наличии анатомических признаков асимметрии все же нет достаточно выраженной функциональной асимметрии, хотя и отмечается незначительное доминирование функций правой гемисферы. Характерно, что у детей в первые два года жизни травмы левой височной доли не ведут к фатальным последствиям, как это имеет место у взрослых, — их речевые функции успешно развиваются в соответствующей зоне правого полушария. Но в более позднем возрасте такое дублирование невозможно, т.к. реализация запускаемых генами развития программ формирования мозга ребенка приводит к окончательному закреплению речевой функции только за левым полушарием.
Экспериментальные данные также показывают, что поражения височных долей неокортекса у шимпанзе не приводят к нарушению вокализации, к каким-либо фатальным изменениям в репертуаре криков и звуков, которые контролируются их лимбической системой. Но, в отличие от шимпанзе, звуковой язык людей управляется неокортексом — это перемещение локализованного центра управления звуковым языком соответствовало важному эволюционному переходу к обучению общению. Зачатки латерализации функций у шимпанзе, их удивительная способность усваивать язык жестов и общаться с человеком указывают на начальные стадии этого перехода. Об этом также свидетельствуют недавно обнаруженные у карликовых шимпанзе (бонобо) простейшие рудименты звуковой речи. Поэтому можно предположить, что начало формирования каких-то зачатков символьного звукового языка у гоминид — это событие весьма отдаленного прошлого, которому несколько миллионов лет. Возможно, первопроходцем здесь оказался Homo habilis, т.к. по останкам черепа представителя этого первого вида гоминид удалось вычислить, что в его мозге уже сформировалась ответственная за речепродукцию зона Брока.
Возникновение Ф. а. м. полностью соответствует принципу биологической (когнитивной) эволюции, согласно которому более высокий уровень организации когнитивных функций влечет за собой их большую дифференциацию между системами. Так как мозг крайне важен для выживания (около 70% ген из генома человека обеспечивают формирование и функционирование нейронных систем его мозга), то дифференциация и функциональная специализация более всего развиты в центральной нервной системе. Если верно, что для первых гоминид уже были характерны зачатки латерализации функций и специализация левого полушария в знаково-символическом мышлении, то вполне естественно, что речь и речевые функции впоследствии оказались привязанными именно к этому полушарию, а его аналитическая стратегия обработки когнитивной информации — к словам как определенным сочетаниям звуковых символов. Развивающаяся дифференциация когнитивных функций человеческого мозга и появившаяся в связи с этим способность к образному и знаково-символическому (логико-вербальному) мышлению в огромной мере увеличили адаптивные возможности гоминид. Благодаря естественному отбору Ф. а. м. закрепилась в виде генетической программы, направляющей формирование обоих когнитивных типов мышления в правом и левом полушарии (порознь), а также их конкретное соотношение (относительное доминирование одного из них).
И. П. Павлов и сотрудники его лаборатории при изучении условно-рефлекторных реакций собак обратили внимание на индивидуальные различия в их проявлении. Эти различия касались скорости и точности образования условных реакций, их интенсивности, способности к адекватному реагированию на изменения раздражителей, общего поведения в экспериментальной ситуации. И. П. Павлов выдвинул гипотезу о том, что в основе этих различий лежат фундаментальные свойства нервной системы, характеризующиеся силой, уравновешенностью и подвижностью процессов возбуждения и торможения. Индивидуальные особенности ВНД проявляются в разной скорости образования и упрочения условных рефлексов, в различной скорости выработки внутреннего торможения, в различной трудности переделки сигнального значения условных раздражителей, в различной работоспособности корковых клеток и т.п. Для каждого индивидуума характерна определенная комбинация основных свойств корковой деятельности. Она и получила название типа ВНД.
Свойства корковых процессов [13]. Представление о типологических особенностях нервной системы человека и животных является одним из определяющих в павловском учении о высшей нервной деятельности. Систематизация типов высшей нервной деятельности основана на оценке трех основных особенностей процессов возбуждения и торможения: силы, уравновешенности и подвижности, выступающих как результат унаследованных и приобретенных индивидуальных качеств нервной системы. Тип как совокупность врожденных и приобретенных свойств нервной системы, определяющих характер взаимодействия организма и среды, проявляется в особенностях функционирования физиологических систем организма и прежде всего самой нервной системы, ее высших «этажей», обеспечивающих высшую нервную деятельность.
Типы высшей нервной деятельности формируются на основе как генотипа, так и фенотипа. Генотип формируется в процессе эволюции под влиянием естественного отбора, обеспечивая развитие наиболее приспособленных к окружающей среде индивидов. Под влиянием реально действующих на протяжении индивидуальной жизни условий внешней среды генотип формирует фенотип организма.
Понятия тип нервной системы и тип BHД чаще всего используются как синонимы, хотя не все ученые с этим согласны, полагая, что с помощью рефлекторных методик в большей степени выявляются особенности функционирования коры большого мозга и в меньшей степени — особенности нервной системы в целом. Однако исследование типов ВНД приближает нас к пониманию истинных типов нервной системы, а значит, дает возможность понять характерологические особенности человека.
Особенности ВНД определяются характером взаимодействия, соотношением основных корковых процессов - возбуждения и торможения. Поэтому в основу классификации типов ВНД положены различия основных свойств этих нервных процессов. Такими свойствами являются:
1. Сила нервных процессов. В зависимости от работоспособности корковых клеток нервные процессы могут быть сильными и слабыми. Сила процесса возбуждения отражает работоспособность нервной клетки. Она проявляется прежде всего в функциональной выносливости, т.е. в способности выдерживать длительное или кратковременное, но сильное возбуждение, не переходя в состояние охранительного торможения. Если слабое раздражение вызывает сильное возбуждение, ведущее к развитию охранительного торможения, то сила процесса возбуждения (работоспособность нервной системы) слабая. Сила процесса торможения понимается как функциональнаяработоспособность нервной системы при реализации условного торможения. Естественно, и безусловное торможение должно быть сильным. Силу процесса торможения следует понимать как свойство нервной системы (аналогично силе процесса возбуждения), а не как текущее состояние. Сила процесса торможения проявляется в способности к образованию различных тормозных условных реакций — угасания, дифференцировки или запаздывания. Сила процесса торможения всегда проявляется там, где имеют место запреты, воздержание от определенных действий или отсрочка реакции. Чем сильнее процесс торможения, тем адекватнее реакция в таких ситуациях.
2. Уравновешенность нервных процессов. В зависимости от соотношения возбуждения и торможения они могут быть уравновешенными или неуравновешенными. Уравновешенность означает равенство силы процессов возбуждения и торможения, вовлеченных в выработку условных рефлексов или условного торможения. В жизни людей часто возникает необходимость торможения сильных возбуждений, чтобы осуществить реакции на другие интенсивные раздражители окружающей среды, и нервные клетки должны выносить эти чрезвычайные напряжения своей деятельности.
3. Подвижность нервных процессов, т.е. быстрота их возникновения и прекращения, легкость перехода от одного процесса к другому. В зависимости от этого нервные процессы могут быть подвижными или инертными.Лабильность, подвижность нервных процессов оцениваются в показателях прочности переделки сигнального значения условных раздражителей (с возбудительного на тормозной и наоборот). Подвижность нервных процессов, согласно представлениям И.П.Павлова, указывает на способность нервной системы быстро реагировать на изменения среды и переходить от одних условных рефлексов к другим в зависимости от требований среды. Подвижность нервных процессов проявляется в способности к изменению поведения в соответствии с изменяющимися стимулами и определяется скоростью возникновения и прекращения нервных процессов
Общие типы ВНД. Теоретически мыслимо 8 комбинаций этих трех свойств нервных процессов. И.П. Павлов, выделил, однако, только 4, наиболее резко бросающихся в глаза типа ВНД у собак (рис. 1):
1 - сильный неуравновешенный (с резким преобладанием возбуждения);
2 - сильный неуравновешенный подвижный;
3 - сильный уравновешенный инертный;
4 - слабый тип.
Связь типов ВНД и темперамента (лат. Tетреramentum — соразмерность, надлежащее соотношение). Древнегреческий врач, основатель медицины Гиппократ (460 —377 до н.э.) объяснял неодинаковое течение одной и той же болезни у разных людей за счет разного соотношения «соков тела» в организме человека: крови, слизи, желчи, черной желчи. Основываясь на этой концепции, Гален выделил четыре темперамента: сангвинический (от лат. sangius — кровь),холерический (от лат. chole — желчь), флегматический (от лат. phlegта — слизь, мокрота), меланхолический (от греч, melanos + chole — черная, испорченная желчь).
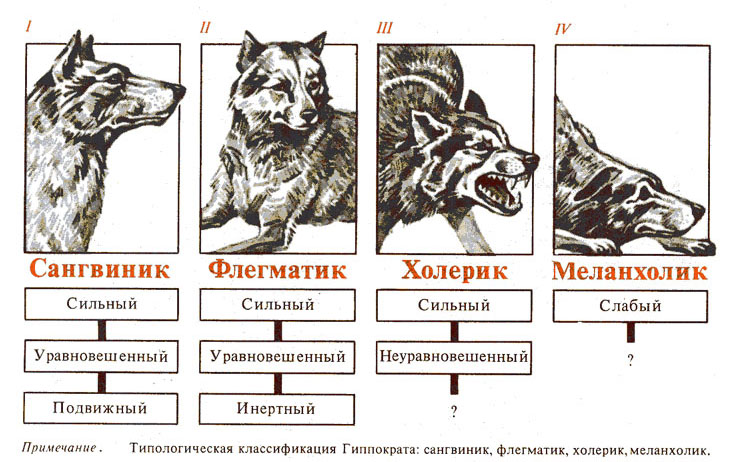
Рис. 1. Типы высшей нервной деятельности собак по И.П. Павлову и их соответствие типам темперамента по Галену (Гиппократу) [ 1 ].
Выделенные типы Павлов считал общими и для человека и животных. Он показал, что четыре установленных типа совпадают с галеновским описанием четырех темпераментов человека - холерическим, сангвиническим, флегматическим и меланхолическим, основанным на гиппократовской классификации основных жидкостей организма.
В формировании типа ВНД наряду с генетическими факторами (генотип) принимает активное участие и внешняя среда, воспитание (фенотип). В ходе дальнейшего индивидуального развития человека на основе врожденных типологических особенностей нервной системы под влиянием внешней среды формируется определенная совокупность свойств ВНД, проявляющаяся в устойчивой направленности поведения, т.е. то, что мы называем характером. Тип ВНД способствует формированию определенных черт характера. По мнению В.М. Смирнова, в определении понятия «тип ВНД» должен присутствовать элемент связи с темпераментом личности. С учетом подобного подхода «тип ВНД – это совокупность врожденных и сформированных в онтогенезе свойств нервной системы (силы, уравновешенности и подвижности процессов возбуждения и торможения), определяющих темперамент личности.»