

26. Коренева система як орган поглинання води. Кореневий тиск. Плач.
Морфологічні та анатомічні особливості будови кореня пов’язані з важливішою його функцією – вбирати з ґрунту воду та мінеральні речовини. На рисунку 5.10 від- творено основні елементи первинної будови кореня: кореневий чохлик, апікальна меристема, ризодерма, первинна кора, ендодерма, перицикл та центральний циліндр, який складається з провідних тканин – ксилеми та флоеми.
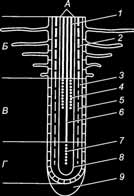
Рис. 5.10. Схема зон та диференціювання тканин кінчика кореня: А – центральний циліндр; Б – зона кореневих волосків; В – зона розтягнення;
Г – зона меристеми; 1 – зрілі елементи ксилеми; 2 – ендодерма з поясками Каспарі; 3 – первинна кора; 4 – незрілі елементи ксилеми; 5 – ендодерма без поясків Каспарі; 6 – зрілі елементи флоеми; 7 – незрілі елементи флоеми; 8 – перицикл; 9 – кореневий чохлик.
Найінтенсивніше вода всмоктується коренем у зоні кореневих волосків. Як було вже зазначено, сумарна ввсмоктувальна поверхня їх величезна. Але слід відзначи- ти, що й епідермальні клітини, що не утворюють волосків, вбирають воду з такою ж швидкістю на одиницю поверхні, як і клітини, що утворюють волоски. Зрозумі- ло, що останні дуже динамічні, вони щільно прилягають до часток ґрунту, а тому й всмоктують основну масу води, що надходить у рослину.
У ділянці кореня вище зони кореневих волосків, де клітини вже опробковують- ся, інтенсивність усмоктування води знижується. З початком вторинного росту коре- ня вбирання вологи ще більш уповільнюється.
Значну роль у вбиранні води відіграє мікориза у рослин, що її утворюють, – вона створює додаткову поглинаючу поверхню, особливо на старих ділянках кореня.
Від поверхні кореня через клітини кори, ендодерму та перицикл вода надходить до судин ксилеми (рис. 5.11). Через клітини кори є два можливих шляхи транспорту- вання води: симпластний через цитоплазму по плазмодесмах і апопластний через клітинні стінки згідно з законами осмосу.
Більш активне радіальне транспортування води у корені відбувається по апо- пласту, оскільки опір клітинних стінок для неї значно нижчий, ніж опір цитоплазми. На рівні ж ендодерми апопластне транспортування неможливе через непроникність поясків Каспарі. Отже, вода може потрапити в ендодерму лише через оболонку та цитоплазму проникних клітин.
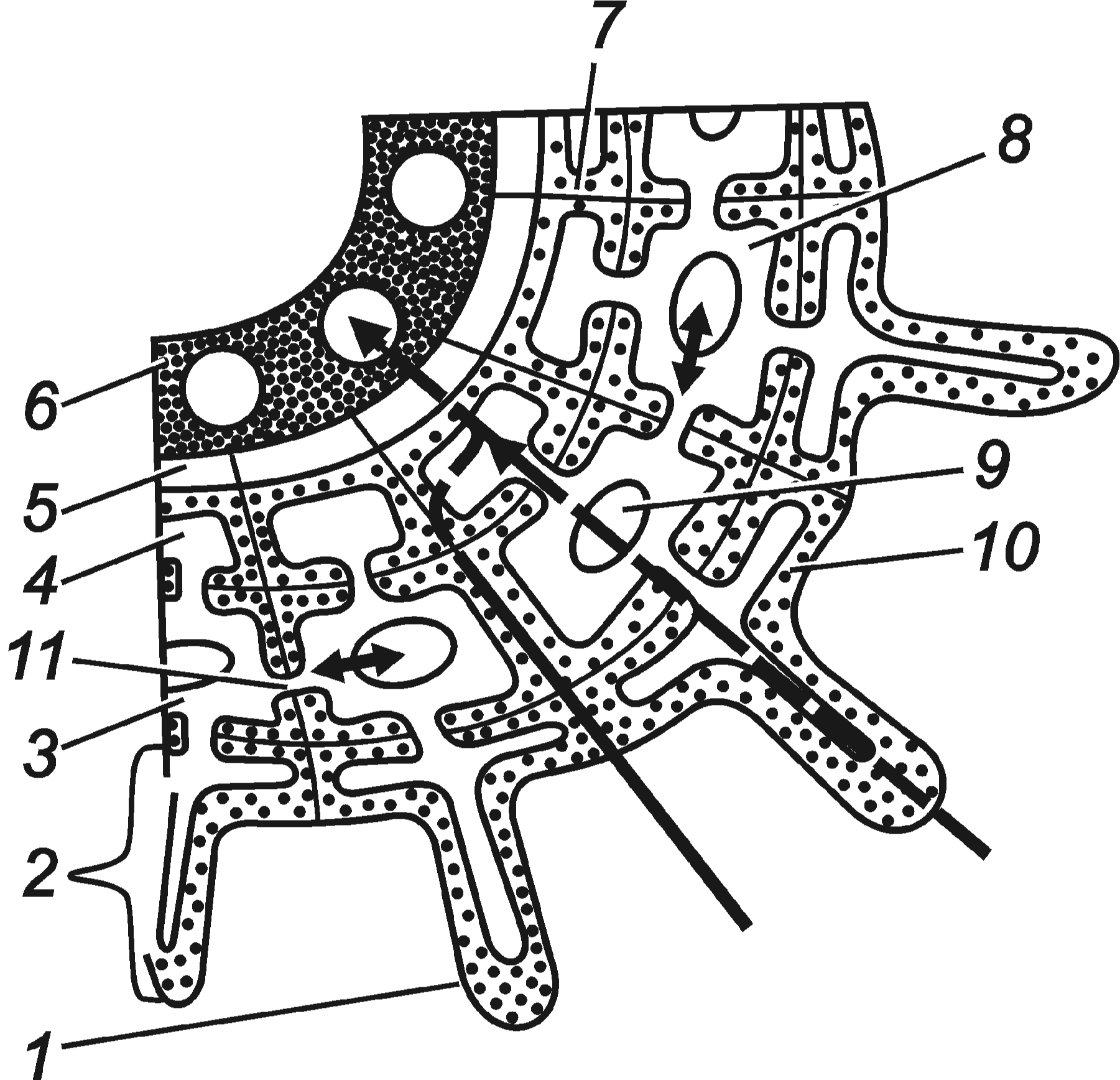
Рис. 5.11. Схема шляхів радіального просування води та іонів через корінь до судин ксилеми: 1 – кореневий волосок; 2 – ризодерма; 3 – паренхіма коры, 4 – ендодерма; 5 – перицикл; 6 – ксилема; 7 – поясок Каспарі; 8 – цитоплазма; 9 – вакуоля, 10 – клітинна стінка; 11 – плазмодесма.
Регулювання току води на рівні ендодерми відбувається внаслідок зміни швид- кого апопластного транспортування на повільний симпластний, а також тому, що діаметр центрального циліндра, куди вона надходить через ендодерму, в 5–6 разів менший діаметра поверхні кори й сисної поверхні кореня. Якщо пояски Каспарі в ро- стучих зонах кореня не повністю формуються, непроникність клітинних стінок для води не є абсолютною. Крім того, в ділянках центрального кореня, де закладаються бокові корінці, ендодерма переривається, що полегшує проникнення води.
Надходження води з ґрунту відбувається за рахунок кореневого тиску, який являє собою силу, що зумовлює направлений рух водного розчину в живих клітинах і виділення його у судини. Механізм підняття води по рослині внаслідок дії корене- вого тиску називається нижнім кінцевим рушієм. Наявність у рослин кореневого тиску, який зумовлює активне поглинання води коренями, добре ілюструється явищами плачу рослин та гутації. Плач рослин – ви- ділення рідини із зрізаної поверхні стебла. Ця рідина називається пасокою. Спочатку виділення її вважали процесом, пов’язаним з пораненням рослин, і що спостерігаєть- ся він лише у деяких рослин у певні пори року. Згодом встановили, що це явище спо- стерігається у всіх рослин й відбувається у будь-якій фазі вегетації, послаблюючись і навіть зникаючи у кінці вегетаційного періоду. Це положення доведено дослідами, у результаті яких установлено, що об’єм виділеної пасоки значно перевищує об’єм кореневої системи. Отже, пасока не може бути рідиною, що витікає внаслідок пора- нення тканин, а результат вбирання кореневою системою навколишнього розчину. Плач рослин поділяють на весняний та літній. Перший відбувається до розпускання листя, другий – після. Весняний плач відрізняється від літнього більшою кількістю соку, що виділяється. Улітку при сильному випаровуванні пасока взагалі не виділяється, а, навпаки, спостерігається від’ємний плач: коли на зрізане стебло налити води, то вона всмоктується пеньком.