

69. Темнова фаза фотосинтезу. Фази карбоксилювання, відновлення, регенерації.
Реакції темнової фази протікають у стромі хлоропласту і не обумовлені наявністю світла безпосередньо. Однак назва “темнова фаза” має більше історичний характер, оскільки, як тепер установлено, світло й у цій фазі використовується для активації ряду ферментів. Цю фазу називають також біохімічною, чи ферментативною, тому що темнові реакції контролюються ферментами.
У вищих рослин відомо три різних механізми темнових реакцій, або вірніше вважати, що є один основний процес і два його варіанти. Основний процес – це фіксація СО2 у циклі Кальвіна, названого так на честь ученого М. Кальвіна (США). Рослини, у яких відбуваються тільки реакції цього циклу, називають С3-рослинами. Вони поширені в географічних зонах з помірним кліматом, оптимальна денна температура для фіксації СО2 у них від 15 до 25 °С. Інші варіанти цього процесу – С4 – шлях і процес, який називають метаболізмом органічних кислот за типом товстянкових. Інакше цей процес називають САМ-метаболізм (від анг. Crassulacean acid metabolism). Рослини- С4, що здійснюють С4-фотосинтез, поширені в зоні посушливого тропічного клімату, але можуть рости й у зонах з помірним кліматом. Багато сукулентів, що живуть у зонах з різко посушливим кліматом, здійснюють САМ-метаболізм. У С4-рослин і САМ-рослин також відбувається цикл Кальвіна, але процес перетворення СО2 у вуглеводи також включає й інші реакції, специфічні для кожної групи рослин.
М. Кальвін зі співробітниками вивчав (1946–1953 рр.) послідовність перетворень вуглецю в реакціях фотосинтезу, використовуючи нові на той час методи. Він працював з радіоактивним довгоживучим ізотопом вуглецю 14С (період напіврозпаду 5720 років), а також використовував радіоавтографію і мало відому в той час паперову хроматографію. Після експозиції 5 с і менше мічений вуглець знайшли в першому продукті фотосинтезу. Цим продуктом виявилася фосфогліцеринова кислота, що має три атоми вуглецю. Звідси інша назва циклу Кальвіна – шлях С3. Уже через одну хвилину після початку інкубації в рослині з 14СО2 синтезувалися цукри, органічні кислоти, різні амінокислоти.
Пізніше Кальвін з’ясував весь ланцюг послідовних ферментативних перетворень цих сполук і виділив у ньому три етапи: карбоксилування, відновлення і регенерацію. Енергія, що забезпечує цикл Кальвіна, надходить у формі АТФ і НАДФ·Н2, що утворилися у світловій фазі.
Вихідним, а також кінцевим продуктом циклу Кальвіна є п’ятивуглецевий цукор із двома фосфатними групами – рибульозо-1,5-бісфосфат (РБФ). Ключова реакція циклу Кальвіна – та, у якій бере участь СО2. Процес карбоксилування починається, коли СО2 входить у цикл і фіксується на РБФ. У результаті утворюється нестійкий проміжний продукт – цукор з 6 атомами вуглецю, що відразу ж розкладається на дві молекули 3-фосфогліцеринової кислоти (ФГК). ФГК і є першим продуктом фотосинтезу.
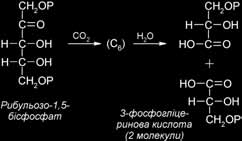
Карбоксилуючим ферментом є рибульозобіфосфаткарбоксилаза (Рубіско). Цей найважливіший фермент знаходиться в стромі хлоропластів у великій кількості: його вміст складає 50 % усіх розчинних білків листка. Вважають, що це найпоширеніший у природі білок. Фермент складається з 8 великих субодиниць, що виконують каталітичні функції, і 8 малих регуляторних субодиниць, організованих у єдину структуру. Для прояву каталітичної активності Рубіско має потребу в присутності іонів Мg2+. Крім карбоксилазної, фермент має також оксигеназну активність, що виявляється в процесі фотодихання. Карбоксилазна чи оксигеназна активність ферменту залежить від концентрації СО2 і О2 у стромі хлоропласта. Субстрати СО2 і О2 взаємно конку- рентні. Як субстрат РуБФ-карбоксилаза використовує переважно СО2.
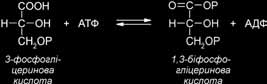
Далі 3-фосфогліцеринова кислота фосфорилюється з утворенням 1,3-біфосфо- гліцеринової кислоти (1,3-БФГК). Фосфорилювання відбувається при участі АТФ, синтезованого у світловій фазі фотосинтезу і ферменту фосфогліцераткінази.
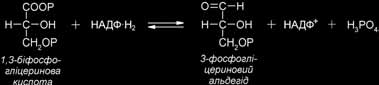
Далі відбувається етап відновлення: 1, 3-БФГК відновлюється до 3-фосфогліцеринового альдегіду (3-ФГА) за участю НАДФ·Н2 продукту світлової фази фото- синтезу. Цю реакцію каталізує фермент гліцеральдегід-3-фосфат-дегідрогеназа.
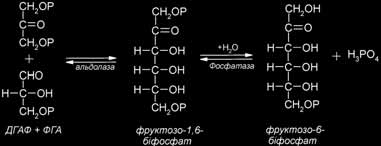
Тріоза 3-ФГА – це перший цукор, що утворюється при фотосинтезі. Як тільки утвориться 3-ФГА власне фотосинтез закінчується, тому що всі подальші реакції відбуваються й у нефотосинтезуючих організмах (тварин, грибів). Молекули 3-ФГА, утворені в циклі Кальвіна, використовуються різними способами: 1/6 їхня частина використовується на синтез вуглеводів, серед яких переважають сахароза і крохмаль, 5/6 – на регенерацію рибульозобіфосфату – первинного акцептора СО2. У випадку, якщо 3-ФГА використовується для синтезу вуглеводів, він ізомеризується в дигідроксиацетонфосфат (ДГАФ). Цю реакцію каталізує фермент тріозофосфатізомераза. Далі дві тріози – 3-ФГА і ДГАФ поєднуються й утворюється цукор фруктозо-1,6-біфосфат. Цю реакцію каталізує фермент фруктозобіфосфа- тальдолаза. Утворений фруктозо-1,6-біфосфат дефосфорилюється з утворенням фруктозо-6-фосфату. Фосфатну групу відщеплює фруктозобіфосфатаза. Фруктозо-6-фосфат виводиться з циклу Кальвіна і за допомогою ферменту глюкозофосфатізомерази перетворюється в глюкозо-6-фосфат. Таким чином, з’являються монофосфати глюкози і фруктози, з яких пізніше утворюється два основних вуглеводи – сахароза і крохмаль.

Для
безперервної фіксації СО2
необхідна
постійна регенерація акцептора СО2
–
рибульозобіфосфату. Тому цикл Кальвіна
замикає фаза регенерації. Регенерація
– це складний багатоетапний процес,
у якому беруть участь цукрофосфати з
3, 4, 5, 6, 7-атомами вуглецю. У результаті
їх перетворень утворюється
рибульозо-5-фосфат. На його фосфорилювання
витрачається інший АТФ, синтезований
у світловій фазі фотосинтезу, і
утворюється рибульозо-1,5-біфосфат, тобто
регенерується вихідна сполука циклу
Кальвіна. По суті, у циклі Кальвіна
рибульозо-1,5-біфосфат піддався
карбоксилюванню, переніс ряд перетворень
і був знову регенерований. Схематично
цикл Кальвіна, який називають також
відновним пен- тозофосфатним шляхом,
можна зобразити так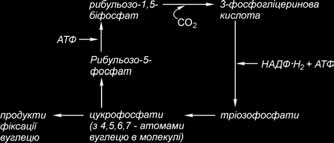
У кожному обороті циклу одна молекула СО2 відновлюється, а молекула рибу- льозобіфосфату регенерується. Для утворення глюкози необхідні 6 обертів циклу з поглинанням 6 атомів вуглецю