

50. Поняття про пігментні системи та реакційний центр.
В даний час встановлено , що фотосинтетичні пігменти в мембранах хлоропластів мають не безладне розташування , а організовані у дві пігментні системи - фотосистему І (ФС I ) і фотосистему II ( ФС II).
Існування двох фотосистем вдалося встановити завдяки тому , що пігменти, що входять до складу ФС я і ФС II , відрізняються за спектральним властивостям . Інтенсивність фотосинтезу при освітленні світлом з довжиною хвилі 680-700 нм може бути значно підвищена додаванням світла з коротшою довжиною хвилі ( 650-660 нм) , і навпаки. Виявилося , що інтенсивність фотосинтезу при висвітленні змішаним світлом ( 650-700 нм) вище суми інтенсивностей фотосинтезу , що спостерігається при освітленні світлом кожного із зазначених вище діапазонів довжин хвиль окремо ( ефект Емерсона ) . Це вказує на те , що у фотосинтезі беруть участь обидві фотосистеми одночасно.
Кожна фотосистема складається з світлозбиральних ( антенних ) молекул пігментів ( хлорофілу а , хлорофілу Ь , каротиноїдів , фікобілінов ) і реакційного центру ( РЦ) . Реакційний центр , в свою чергу , включає фотоактивний пігмент - пастку і первинні донори та акцептори електронів. Пігмент- пастка ФС І поглинає світло з довжиною хвилі 700 нм і позначається Р700 (або П700 ) , а пігмент -пастка ФС II поглинає світло з довжиною хвилі 680 нм і позначається Р680 (або П680 ) .
Пігменти антенного комплексу поглинають світло в тій частині спектра , в якій не поглинає пігмент -пастка , і доставляють поглинену енергію в РЦ , що дозволяє ефективніше використовувати енергію світла . Як правило , вони поглинають світло , з довжиною хвилі меншою , ніж світло поглинається хлорофілом , що входять в РЦ . Перенесення енергії відбувається тільки від пігментів , що поглинають світло з меншою довжиною хвилі , до пігментів , що поглинає світло з більшою довжиною хвилі.
Справа в тому , що хоча передача енергії від однієї молекули пігменту до іншої йде з великою ефективністю (від хлорофілу б до хлорофілу а - 90 %, від каротиноїдів до хлорофілу - 40 %) , однак все ж це пов'язано з деякою втратою енергії. Втрата енергії призводить до перетворення квантів з більшою енергією ( з меншою довжиною хвилі) в кванти з меншою енергії ( з більшою довжиною хвилі). Саме тому основні форми хлорофілу , до якого стікається енергія , є найбільш « довгохвильовими ». Зворотній перенос енергії неможливий.
Треба відзначити , що функції білково- пігментного антенного комплексу полягають не тільки в поглинанні і передачі енергії , але і в захисті хлорофілу від активних форм кисню , які утворюються при поглинанні світла

Рис. 1. Спрощена модель фотосистеми.
І. Антенний комплекс. II. Реакційний центр.
1. Кванти світла. 2. Пігменти, що поглинають в короткохвильовій частині спектра видимого світла. 3. Пігменти, що поглинають в средньохвильовій частини спектра видимого світла. 4. Молекули хлролофіла з різними спектрами поглинання. 5. Хлорофіл-пастка P680 або P700. 6. Шлях енергії поглиненого кванта світла до хлорофілу-пастки.
Крім світлозбирального пігмент-білкового комплексу і комплексів ФС I і ФС II в фотосинтезуючих мембранах знаходиться так званий цитохромний комплекс, що забезпечує перенесення електронів між фотосистемами і циклічний перенос електронів навколо ФС I, а також рухомі переносники електрона (пластохінон, пластоціанін, фередоксин).
62. С4 – шлях фотосинтезу
С4 – шлях фотосинтезу був виявлений у 50–60 р. ХХ ст. у двох тропічних злаків (кукурудзи і цукрової тростини), а пізніше й у багатьох інших рослин. У 1965 р. Ю. С. Карпілов із співробітниками знайшли продукти темнової фази фотосинтезу, що відрізнялися від циклу Кальвіна. У короткострокових дослідах з 14С у кукурудзи мічений вуглець знаходили в складі незвичних продуктів фотосинтезу – органічних кислот з 4 вуглецевими атомами – щавлевооцтової, яблучної й аспарагінової. Аналогічні результати були отримані в дослідах із цукровою тростиною. У 1966 р. австралійські вчені М. Хетч і К. Слек описали шлях С4 як новий тип асиміляції СО2, відмінний від циклу Кальвіна. У їх честь С4-шлях фотосинтезу називають та- кож циклом Хетч – Слека.
Вони встановили, що шлях С4 включає дві реакції карбоксилювання і що ці реакції розділені в просторі. Специфічність механізму фотосинтезу обумовлена особливостями фізіології й анатомічної будівлі листків С4-рослин. Для листя цих рослин характерні численні повітряні порожнини, по яких СО2 з атмосфери безпо- середньо проникає до дуже великої кількості фотосинтезуючих клітин. У рослин С4 навколо провідного пучка лежать щільно упаковані великі паренхімні клітини обкладки, утворюючи ніби корону. Навколо них менш щільними шарами розміща- ються невеликі клітини мезофілу.
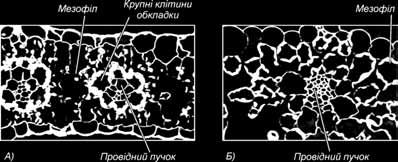
Анатомічні відміни поперечного зрізу листа між С4 та С3 рослинами (370Х): А – рослина Zea mays; Б – рослина Avena Sativa.
Хлоропласти клітин обкладки і клітин мезофілу різні за структурою: у хло- ропластах клітин обкладки зовсім немає гран або вони дуже малі. Хлоропласти клітин мезофілу мають добре розвинуті грани. У хлоропластах клітин мезофілу накопичується дуже мало крохмалю, а в хлоропластах клітин обкладки утворю- ються численні великі крохмальні зерна. Клітини мезофілу й обкладки з’єднані великою кількістю плазмодесм.
На противагу циклу Кальвіна в клітинах мезофілу С4-рослин акцептором СО2 (у формі НСО3) виступає тривуглецева сполука – фосфоенолпіровиноградна кис- лота – ФЕП (фосфоенолпіруват). Тому першим продуктом асиміляції є чотири- вуглецева сполука – щавлевооцтова кислота (ЩОК, оксалоацетат). Звідси і назва циклу – С4. Цю реакцію каталізує фермент фосфоенолпіруваткарбоксилаза (ФЕП- карбоксилаза) у присутності іонів магнію, що знаходиться не в хлоропластах, як рибульозобісфосфаткарбоксилаза, а в цитоплазмі клітин мезофілу. Таким чином, оксалоацетат утворюється в цитоплазмі. ФЕП-карбоксилаза має високу спорідненість до СО2, тому фотосинтез може відбуватися при майже закритих продихах, що характерно для рослин жаркого клімату.