

7. Нециклічний транспорт електронів.
Розрізняють нециклічний шлях транспорту електронів і циклічний, або нециклічне і циклічне фотофосфорилування. Шлях електронів від води до НАДФ+ у мембранах хлоропластів називають нециклічним потоком електронів, він спо- лучений з нециклічним фотофосфорилуванням. Назва “нециклічний” указує на те, що шлях електронів не замкнутий, нециклічний. Назва “фото” указує на те, що рух електронів відбувається за рахунок енергії світла, а “фосфорилування” пов’язане з тим, що перенос електронів поєднаний із синтезом АТФ з АДФ і ортофосфату.
Нециклічне фотофосфорилування зображують сумарним рівнянням:
![]()
14. Еволюція фотосинтезу. Фоторедукція.
Первинні , здатні до самовідтворення живі утворення виникли в результаті абіогенної хімічної еволюції(згідно загальновизнаної нині еволюційної теорії походження і розвитку життя , яка більше 50 років тому була сформульована А. І. Опаріним). Будучи оточеними близькими за складом , але ще неживими органічними сполуками , ці первинні істоти могли здійснювати в безкисневому середовищі анаеробний гетеротрофний тип живлення за допомогою невеликого набору ферментів. Поступове виснаження і деградація органічних речовин , синтезованих абіогенним шляхом , супроводжувалося нагромадженням усе більш окиснених сполук , аж до появи найбільш бідного енергією з'єднання вуглецю - вуглекислоти. Це тягло за собою необхідність все більшого і більшого вдосконалення й ускладнення ферментативного апарату , необхідного для асиміляції усе більш окинених речовин. У цих умовах , які все ще характеризувалися відсутністю в середовищі кисню , цілком ймовірне виникнення первинних автотрофних організмів , які здійснювали відновлення вуглекислоти за рахунок хімічної енергії , отриманої з мінеральних речовин. Такий тип живлення отримав назву хеморедукціі .
Поява в цей період , який характеризувався сильно відновними умовами середовища призвело, очевидно , до заміни хімічної енергії в процесах хеморедукціі на світлову . Виник найпростіший тип фотоавтотрофного живлення, який отримав назву фоторедукціі і бактеріального фотосинтезу. Такий тип живлення здійснюють сучасні фототрофні бактерії - пурпурні серобактерии ( Thiorhodaceae ) і зелені серобактерии ( Chlorobacteriaceae ) , у яких роль пігменту - фотосенсибілізатора виконує бактеріохлорофіл і які є облігатними анаеробами . Пурпурні і зелені серобактерии відновлюють вуглекислоту за рахунок енергії світла , використовуючи як Н- донора сірководень (H2S) :
15. Енергетика фотосинтезу.
19. Залежність фотосинтезу від умов оточуючого середовища(світла, со2, температури, кисню).
Світло, промениста енергія – найважливіший з факторів, що визначає інтен- сивність фотосинтезу. У процесі фотосинтезу використовується частина сонячного випромінювання – промені з довжиною хвилі від 380 до 750 нм, так звана фотосин- тетично-активна радіація (ФАР). Умови освітлення рослин включають як інтен- сивність, так і спектральний склад сонячного світла.
К. А. Тімірязєв провів більше 200 дослідів з вивчення світлової залежності швидкості фотосинтезу. Він установив, що з наростанням інтенсивності освітлення швидкість фотосинтезу спочатку стрімко зростає, потім росте усе повільніше і по- вільніше, досягає максимуму і стає стабільною. Іншими словами, світлова крива має вигляд логарифмічної кривої (рис. 3.18).
Занадто висока інтенсивність світла гнітить біосинтез пігментів, ростові проце- си. У С3-рослин насичення світлом відбувається при нижчій його інтенсивності, ніж у С4-рослин. У С4-рослин висока швидкість фотосинтезу спостерігається в основно- му при високому рівні освітленості.
Стосовно оптимуму освітленості рослини поділяють на світлолюбні і тіньо- витривалі. Якщо у світлолюбних світлове насичення настає при освітленості 100 – 300 · 103 ерг/см2/с, то в тіньовитривалих – при освітленості набагато нижчій.
Екологічні групи рослин добре адаптовані до світлового режиму місць проживан- ня. Адаптація досягається шляхом значної зміни структури і функції листя, кількості і співвідношення пігментів, кількості карбоксилуючих ферментів і компонентів елек- тронотранспортного ланцюга, розмірів антенного комплексу. У світлолюбних рос- лин палісадна паренхіма часто буває багатошаровою, хлоропласти дрібні (рис. 3.19). Число продихів на одиницю поверхні велике, що полегшує надходження вуглекис- лоти усередину листа. У світлолюбних рослин зменшений вміст зелених пігментів і збільшений вміст каротиноїдів. Відношення “хлорофіл а: хлорофіл b” у світлолюб- них рослин вище, ніж у затінкових, тобто пристосування до високої інтенсивності світла пов’язане зі зменшенням окислених форм пігментів: хлорофілу b і ксантофілу. У тіньовитривалих рослин збільшується розмір листків і хлоропластів, підвищуєть- ся вміст хлорофілу, що дозволяє їм активніше використовувати низькі інтенсивності світла. У природних умовах верхні і найбільш виступаючі листки мають характер світлових, а листки біля основи гілки за своєю анатомією відповідають затінковим.
Світлолюбні і тіньовитривалі рослини розрізняються за своїм відношенням до точки світлової компенсації. Точка світлової компенсації – це рівень освітлення, при якому поглинання СО2 у процесі фотосинтезу врівноважується виділенням О2 у процесі дихання в тих же самих листках (при 20 °С і 0, 03 % СО2). Світлолюбні рослини мають досить високу інтенсивність дихання, їх компенсаційна точка складає 3–5 % повного сонячного освітлення, а у тіньовитривалих – приблизно 1 % від повного освітлення.
Точка світлової компенсації залежить також від концентрації СО2 у повітрі: при підвищенні концентрації СО2 вона зрушується у бік вищих значень освітленості. Подібне спостерігається і при підвищенні температури. Розташування точки світло- вої компенсації має важливе значення для продуктивності фотосинтезу.
Ще К. А. Тімірязєв установив, що для фотосинтезу, крім інтенсивності освіт- лення, істотний також спектральний склад сонячного світла. Установлено, що най- більш ефективні в процесі фотосинтезу рослин червоні і сині промені. Світлолюбні рослини інтенсивніше використовують червоні, а адаптовані до розсіяного світла рослини – синьо-фіолетові промені. Рослини, вирощені при червоному і синьому світлі, істотно відрізняються за складом продуктів фотосинтезу. За даними дослід- ників, під впливом синіх променів у продуктах фотосинтезу переважають речовини невуглеводної природи – амінокислоти, білки, органічні кислоти; червоні ж промені сприяють утворенню і нагромадженню вуглеводів (сахароза та ін.).
Фотосинтез і концентрація вуглекислоти
Щорічно рослинність нашої планети фіксує 2 · 1011 т С у вигляді вуглекис- лоти. Концентрація вуглекислоти в атмосфері порівняно невелика – 0,03–0,04 % і підтримується майже незмінною за рахунок буферних властивостей океанів і постійного руху повітряних мас.
Вважають, що диоксид вуглецю атмосфери на 90 % має ґрунтове походження. Процеси дихання і розкладання, що безупинно протікають у ґрунті, поповнюють атмосферні запаси СО2. Інтенсивний обмін СО2 між атмосферою і ґрунтом приво- дить до добових і сезонних коливань концентрації СО2 у повітрі: удень за рахунок фотосинтезу його вміст у повітрі падає, уночі за рахунок дихання підвищується. У густих рослинних угрупуваннях (посіви, ліси) добові коливання у повітрі можуть досягати 25 % від середньої величини. Сезонні коливання концентрації СО2 у по- вітрі менш істотні, ніж добові.
Поряд з освітленістю низька концентрація СО2 у повітрі може бути чинником, що обмежує фотосинтез.
Залежність фотосинтезу від вмісту вуглекислоти в повітрі має вигляд логариф- мічної кривої (рис. 2).
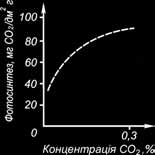
Рис. 2. Інтенсивність фотосинтезу в залежності від концентрації СО2 у люпину.
Фотосинтез і концентрація кисню в атмосфері
Фотосинтез вищих рослин відбувається в аеробних умовах, при концентрації кисню в атмосфері близько 21 %. Така концентрація кисню вища оптимальної, і зни- ження концентрації до 3 % не відбивається на інтенсивності фотосинтезу, а в ряді випадків у рослин з активним фотодиханням (боби та ін.) виявляється сприятливим.
Дуже низька концентрація кисню в середовищі пригнічує фотосинтез, тому що його присутність необхідна для регулювання роботи електронотранспортного ланцюга. Високі концентрації О2 (25–30 %) помітно знижують фотосинтез. Причиною служить інгібуючий вплив високих концентрацій кисню на ключовий фермент фотосинтезу – РуБФ-карбоксилазу-оксигеназу. При зменшенні концентрації СО2 і підви- щеному парціальному тиску О2 пригнічується карбоксилазна функція й активується оксигеназна функція РуБФ-карбоксилази, стимулюється фотодихання.
Фотосинтез і температура повітря
Залежність фотосинтезу від температури виявляється перш за все у темнових ферментативних реакціях, зв’язаних з відновленням СО2, та в процесах фотосинтетичного фосфорилювання. Залежать від температури також і реакції електронного транспорту. Однак фотофізичні реакції, що протікають в реакційних центрах, від температури не залежать. Загальна залежність фотосинтезу від температури має вигляд одновершинної кривої, на якій розташовані кардинальні (основні) точки: мінімум, оптимум та максимум .
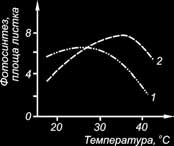
Рис. Температурні оптимуми С3 – (1) и С4 (2) рослин (за А. Т. Макроносовим, В. Ф. Гавриленко).
Мінімум характеризується тією температурою, при якій фотосинтез починається. У більшості рослин помірної зони фотосинтез припиняється приблизно при 0 °С. Однак у деяких хвойних (ялина, сосна звичайна) він спостерігається і при –5, –3 °С. У озимих, альпійських і рослин, що квітнуть навесні, поглинання СО2 спо- стерігається при –7 °С, а в мохів, лишайників холодних областей – при –15 °С. У такий спосіб озимі зелені рослини й узимку ведуть фотосинтез.
При підвищенні температури швидкість фотосинтезу збільшується у 2–3 рази.
Оптимум – це та температура, при якій фотосинтез найстійкіший і має найвищу швидкість. Оптимальні температури лежать у широкому інтервалі: у С4-рослин жарких областей – при 35–45 °С, у сільськогосподарських С3-рослин – при 20–30 °С. Температурний оптимум фотосинтезу строго не фіксований, залежить від стадії розвитку рослин і від даного комплексу зовнішніх факторів.
Максимум – це та температура, після досягнення якої фотосинтез припиняється. Максимуми знаходяться в широкому діапазоні температур між 35° і 50 °С. У більшості С3-рослин при 35° відзначається гальмування фотосинтезу. У С4-рослин жарких областей верхня межа температури для поглинання СО2 – 50– 60 °С. Підвищення температури робить негативним вплив на фотосинтез С3-рослин, тому що при цих умовах зменшується розчинність СО2, збільшується відношення О2/СО2.
У помірному кліматі в теплі місяці температура як фактор фотосинтезу особливої ролі не грає. У польових умовах продуктивність фотосинтезу у границях температури від 16 до 29 °С майже не змінюється. При високих температурах загальна інтенсивність фотосинтезу С4-рослин вища, ніж у С3-рослин.