

66. Стадійний розвиток рослин. Світлова стадія. Явище фотоперіодизму.
Фотоперіодизм – це здатність рослин переходити до цвітіння тільки при певному співвідношенні довжини темного і світлого періоду доби. Він виражається у зміні процесів росту і розвитку, що забезпечують адаптацію онтогенезу конкретного виду рослин до сезонних особливостей кліматичних умов у даному місці його життя. Довжина дня і ночі використовується рослиною як астрономічний годинник, що показує кращий час переходу до активного цвітіння, розвитку бульб і цибулин чи підготовки до сезонних несприятливих умов. У процесі еволюції сформувалися три основні групи рослин з різною фотоперіодичною реакцією: довгоденні, короткоденні, нейтральні.
Довгоденні рослини (ДДР) – жито, ячмінь, пшениця й ін. – вимагають для свого розвитку довгого дня і короткої ночі, зацвітають при довжині дня більше певної (критичної) тривалості.
Короткоденні рослини (КДР) – просо, соя, рис і ін. – вимагають для свого роз- витку довгої ночі і короткого дня, зацвітають при довжині дня менше певної (кри- тичної) тривалості.
Нейтральні рослини – томат, деякі сорти бавовнику й ін. – зацвітають при будь- якій довжині дня.
Більшість рослин довгого дня вирощується в районах помірного клімату. Рослини короткого дня, південного, тропічного походження не переходять до цвітіння в умовах довгого дня цих зон. Це важливо при видовому і сортовому районуванні сільськогосподарських рослин. У різних видів і сортів рослин мінімальний фото- періодичний індуктивний період складає від 1 до 25 діб (циклів) і приводить до на- ступного зацвітання рослин незалежно від довжини дня. Для короткоденних рослин необхідний безперервний темновий період певної довжини. Якщо його перервати коротким (10 хв.) світлом, то індукції цвітіння не буде. Для довгоденних рослин по- трібний довгий світловий період, що прискорює їхній розвиток. Індукція виклика- ється світлом дуже малої інтенсивності (3–5 лк).
Фотоперіодична реакція рослин найбільш успішно проходить у променях певних довжин хвиль, що збігаються зі спектром дії фітохрому. В індукції цвітіння бере участь фітохром (система ФЧ~ ФДЧ), який відповідає за сприйняття співвідношення світла і темряви. Вважається, що білок фіто- хрому це фермент, що каталізує синтез якоїсь фізіологічно активної речовини. У нічний період Фдч перетворюється у Фч і це сприяє початку реакцій, що приводять короткоденні рослини до цвітіння. При спалаху червоного світла (660 нм) Фч перетворюється у Фдч, що гальмує реакції, що приводять до цвітіння.
Якщо рослину короткого дня, наприклад, хризантему, вирощувати на довгому дні, вона не зацвітає. Однак рослина зацвітає при тимчасовому затіненні тільки листя світлонепроникною тканиною. Отже, сприймають фотоперіодичний вплив листки, у яких наявний фітохром. У листках під дією відповідного фотоперіоду проходять процеси, що приводять до утворення особливих речовин, які пересуваються по стеблу і приводять до закладення в конусі наростання квіткових зачатків.
Яка ж природа виникаючого в листках стимулу цвітіння? М. X. Чайлахян (1937) висунув гіпотезу двокомпонентної гормональної системи зацвітання (флориген) (рис. 7.15).
Передбачається, що в гормональний комплекс флоригену входять гібереліни і гіпотетичні антезини. Цвітіння можливе лише при наявності і сприятливому співвідношенні у рослині двох компонентів флоригену – гібереліну й антезину. У довгоденних рослин утворення квіткових стебел залежить від наявності гібереліну, що накопичується в достатній кількості лише на довгому дні. Другий гормон цвітіння – антезин – у довгоденних рослин завжди присутній у достатній кількості.
Варто згадати і про деякі аспекти практичного використання фотоперіодичної реакції при вирощуванні культурних рослин. Змінюючи довжину світло-темнового періоду у декоративних рослин, які вирощують у захищеному ґрунті, можна регулювати цвітіння й одержувати квіти до будь-якого строку, а також збирати кілька врожаїв овочевих протягом року. Вирощування ДДР на короткому дні у вегетаційних будиночках з перериванням темряви спалахом світла дозволить не тільки стимулювати цвітіння рослин і одержувати гарний врожай, але і значно знизити собівартість продукції за рахунок економії електроенергії.
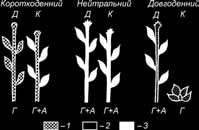
Рис. 7.15. Автономний і індукований механізми регуляції цвітіння нейтральних, короткоденних і довгоденних видів (М. X. Чайлахян, 1989):
1 – гібереліни – гормони, необхідні для утворення стебел; 2 – антезини – гормони, необхідні для утворення квіток; 3 – флориген – гібереліни + антезини, необхідні для цвітіння
У відкритому ґрунті можна зібрати кілька врожаїв ДДР (редис, редька та ін.), якщо в ранній період і під час формування коренеплодів закрити рослини ящиками чи темною плівкою, створюючи їм 12-годинний день. У суниці після одержання навесні першого врожаю можна викликати нове формування квіток, якщо на якийсь час закривати їх, тому що для формування квіток їм необхідний короткий день. Потім рослини знову відкривають і вони дають плоди вже на довгому дні. У цих умовах можна зібрати два врожаї за сезон.