

2.2.2 Электрические синапсы
Электрические синапсы были открыты в 1959 году Фершпаном и Поттером в брюшном нерве краба, а позднее их обнаружили в многочисленных организмах (Рис.26). Передача электрических сигналов в электрических синапсах осуществляется в специализированных структурах – щелевых контактах, где клетки находятся на расстоянии 2 нм и соединяются проводящими каналами (коннексоны). В противоположностьхимическому синапсу, где прохождение импульса несколько задерживается из-за высвобождения и диффузии медиатора, сигнал через электрический импульс передается быстро.
Рис. 26. Электрический синапс.
Электрические синапсы сравнительно редки, и их роль в центральной нервной системе высших организмов неясна. Вместе с тем физиологическая важность электрических синапсов может быть связана с необходимостью быстрой передачи информации.
2.3. Возбуждение и торможение в центральной нервной системе.
Возбуждение и торможение в ЦНС обеспечивается функционированием возбуждающих и тормозных химических синапсов. В возбуждающих синапсах осуществляется перенос нервного импульса от одной клетки к другой. Примерами являются возбуждающее действие центральных отростков первичных афферентых нейронов, мотонейронов и вставочных клеток спинного мозга.
Для возбуждения нейрона необходимы поток афферентных импульсов и их взаимодействие. Один, пришедший к нейрону возбуждающий сигнал, сопровождается развитием локальной деполяризации на постсинаптической мембране, обозначаемой как возбуждающий постсинаптический потенциал (ВПСП). Возникновение ВПСП связано с одновременным увеличением проницаемости постсинаптической мембраны для Nа+ и К+, а также, возможно, Са++. Амплитуда ВПСП составляет всего 0,05 мв. Пороговый же потенциал, необходимый для генерации ПД составляет – 5 – 10 мв. Поэтому для возбуждения нейрона требуется множество импульсов. Наиболее эффективны синаптические контакты на теле нейрона, т.к. постсинаптические мембраны этих синапсов располагаются в непосредственной близости от аксонного холмика, места первичного возникновения ПД (генераторный пункт нейрона). Отличительной особенностью мембраны аксонного холмика является ее высокая возбудимость, отсутствие синапсов и миелиновой оболочки, высокая концентрация Nа – каналов. ВПСП электротонически достигает аксонного холмика, обеспечивая здесь уменьшение мембранного потенциала до критического уровня. В этот момент в аксонном холмике возникает ПД, который ортодромно переходит на аксон и антидромно на тело нейрона. Деполяризация нейрона при действии возбуждающего медиатора может быть достигнуто не только за счет увеличения проницаемости для Nа+ (или Са++), но и за счет уменьшения проницаемости для ионов К+.
Дискуссионным остается вопрос о роли дендритов в возникновении возбуждения. Дендритные синапсы удалены на значительное расстояние от генераторного пункта нейрона. В этой связи, возникающие на постсинаптической мембране дендритов ВПСП, не могут вызвать там должную деполяризацию для генерации ПД. Считают, что синаптический аппарат дендритов проявляет себя при одновременном поступлении сигналов к значительному числу дендритных синапсов. При этом суммарный дендритный ВПСП может изменить мембранный потенциал генераторного пункта на подпороговый уровень и вызвать лишь модуляцию его возбудимости, делая ее большей или меньшей в зависимости от временных и амплитудных характеристик колебаний мембранного потенциала генераторного пункта относительно величины критического уровня деполяризации. Это может сказаться на хараетере ответной реакции нейрона при поступлении к нему в этот момент возбуждений через аксосоматические синапсы. Подобный способ синаптического возбуждения нейрона очень важен для его интегративной функции, т.е. способности суммировать влияния, поступающие на нейрон по разным синаптическим путям.
Афферентные волокна по своему действию являются чисто возбуждающими. Но в центральной нервной системе пришедшее по ним возбуждение направляется по сложным проводящим путям, которые могут быть возбуждающими и тормозными. Последние определяется характером тех вставочных нейронов, которые составляют этот проводящий путь.
Впервые торможение в ЦНС открыл И.М. Сеченов в 1863 году. В опыте на таламической лягушке он показал, что латентное время сгибательного рефлекса при погружении задней конечности в слабый раствор серной кислоты, значительно удлиняется, если на зрительный бугор предварительно положить кристаллик поваренной соли. Это открытие послужило толчком для дальнейших исследований торможения в ЦНС. Под торможением в ЦНС понимают активный нервный процесс, результатом которого является прекращение или ослабление возбуждения в ЦНС В настоящее время общепризнанно существование тормозных интернейронов. Пресинаптические окончания аксонов тормозных нейронов образуют тормозные синапсы. В тормозных синапсах полученный клеткой импульс сопровождается подавлением другого возбуждения. Примером тормозных нейронов в спинном мозге являются вставочные нейроны Реншоу, в головном мозге - грушевидные нейроны Пуркинье коры мозжечка. Обнаружено два совершенно различных механизма торможения: пресинаптический и постсинаптический.
Пресинаптическое торможение. В случае пресинаптического торможения вставочные тормозные нейроны образуют синапсы на возбуждающих пресинаптических окончаниях (аксо-аксональные синапсы). Тормозный медиатор (ГАМК), выделяемый вставочными нейронами, деполяризует мембрану пресинаптического окончания. Это связано с повышением проницаемости мембраны пресинаптических терминалей для ионов Сl-, которые начинают выходить наружу согласно электрическому градиенту. В очаге деполяризации нарушается процесс распространения возбуждения, в результате чего уменьшается или прекращается выделение пресинаптическими окончаниями афферентного нейрона возбуждающего медиатора. Это, в свою очередь, проявляется в подавлении ВПСП и, следовательно, в прекращении поступления на мотонейрон возбуждающих сигналов по определенным афферентным волокнам.
Таким образом, пресинаптическое торможение ограничивает и регулирует афферентацию мотонейрона. Вставочные нейроны с тормозным пресинаптическим действием обнаружены в спинном мозгу, а их синапсы на окончаниях первичных афферентных волокнах. (Рис. 27).
Рис. 27.
Деполяризация пресинаптических терминалей может возникнуть также при накоплении ионов К+ в межклеточной жидкости в результате повышенной активности нервных окончаний и соседних нервных клеток.
Постсинаптическое торможение. Этот вид торможения имеет особенно широкое распространение в ЦНС. Вставочные нейроны образуют тормозные синапсы на дендритах и телах мотонейронов, нервных клетках мозгового ствола, гиппокампа и коры. Выделяемый здесь медиатор (глицин или гамма – аминомасляная кислота) гиперполяризует постсинаптическую мембрану, т.е. увеличивает мембранный потенциал этой области. Эта гиперполяризация получила название тормозного постсинаптического потенциала (ТПСП) Временное течение ТПСП почти совпадает с временным течением ВПСП (Рис.28).
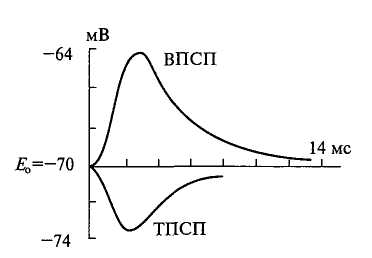
Рис. 28. Возбуждающий (ВПСП)
и тормозной (ТПСП) постсинапти-
ческие потенциалы
Действие тормозного медиатора при постсинаптическом торможении связано с увеличением проницаемости постсинаптической мембраны мотонейрона для ионов калия и хлора. Действие глицина на ионотропный рецептор постсинаптической мембраны сопровождается увеличением ее проницаемости для ионов хлора. Ионы хлора поступают в клетку согласно концентрационному градиенту, в результате чего развивается ТПСП. В безхлорной среде тормозная функция глицина не реализуется.
При действии гамма – аминомасляной кислоты на постсинаптическую мембрану ТПСП развиваетсяв результате входа ионов хлора в клетку или выхода ионов калия из клетки.
Абсолютная величина ТПСП на 10 мв больше мембранного потенциала мотонейрона в покое. Эффективность тормозных синапсов во многом зависит от их локализации на поверхности клетки. Тормозный эффект тем более значителен, чем ближе тормозный синапс расположен к месту генерации потенциала действия. Вследствие этого тормозные синапсы локализованы главным образом на теле нервных клеток вблизи триггерной зоны аксонного холмика.
При активации нескольких тормозных синапсов происходит суммация ТПСП и мембранный потенциал всего мотонейрона увеличивается. Гиперполяризация мотонейрона может развиваться и вследствие временной суммации ТПСП.
Если на фоне гиперполяризации мотонейрона, вызванной суммацией ТПСП к клетке приходят возбуждения и возникают ВПСП, то деполяризующее действие ВПСП может оказаться недостаточным для достижения критического уровня деполяризации мотонейрона и его возбуждения. Таким образом, конечный эффект - возбуждение или торможение будет зависеть от преобладающего действия возбуждающих или тормозных синапсов.
Существует несколько разновидностей постсинаптического торможения: прямое, или реципрокное торможение и возвратное, или антидромное торможение в спинном мозгу.
Понятие реципрокного торможения было предложено Шерригтоном для объяснения работы мышц - антагонистов. Современное представление об этом способе постсинаптического торможения было разработано Экклсом (1952 г.)
Импульсы, идущие по афферентным волокнам, отходящих от мышечных веретен (рецепторов растяжения), возбуждают рецепторы своей мышцы (т.е. мышцы, от которой отходят афферентные волокна) и одновременно через тормозные вставочные нейроны тормозят мотонейроны мышцы – антагониста. Такие же отношения создаются между одноименными мышцами симметричных конечностей. (Рис.29).
Рис. 29. А - прямой тормозный путь и реципрокные отношения между мышцами - сгибателями и разгибателями; Б - схема возвратного торможения.
Р- мышца -разгибатель голени,
С - мышца - сгибатель,
М - мотонейрон,
ТН- тормозные нейроны,
кл. Р - клетки Реншоу.
Возвратное постсинаптическое торможение осуществляется клетками Реншоу (тормозный медиатор – глицин). Аксоны мотонейронов, идущие к мышцам, дают возвратную веточку в спинном мозгу, которая оканчивается на клетках Реншоу. Аксон клетки Реншоу образует тормозные синапсы на теле мотонейрона. Таким образом, мотонейрон при своем возбуждении стимулирует клетку Реншоу, которая тормозит этот же мотонейрон. Это торможение в центрах мышц-сгибателей и разгибателей обеспечивает поочередное сокращение и расслабление скелетной мышцы, что необходимо при ходьбе и беге.
Постсинаптическое торможение мотонейронов спинного мозга может быть вызвано нисходящими влияниями от различных уровней головного мозга (кора больших полушарий, средний мозг, мозжечок и др.). Примером такого вида постсинаптического торможения спинномозговых рефлексов может служить феномен Сеченовского торможения. Стимуляция клеток ретикулярной формации промежуточного мозга приводит к возбуждению тормозных вставочных нейронов спинного мозга и торможению мотонейронов.
Торможение является важным фактором, обеспечивающим координационную деятельность ЦНС, играет важную роль в обработке поступающей в ЦНС информации, выполняет охранительную роль. Блокада торможения ведет к широкой иррадиации возбуждения и судорогам, например при выключении пресинаптического торможения бикукулином, антагонистом ГАМК.