

Коэффициенты корреляции рангов между абсолютными порогами зрения, порогами при электрической адекватной стимуляции и силой нервной системы по данным четырех методик (в.Д. Небылицын, 1960в)
|
Угашение с подкреплением |
«Утомление» |
«Повторение» |
«Форма кривой» |
Реобаза |
59* |
74*** |
|
54* |
Абсолютный зрительный порог |
68** |
44 |
51* |
53* |
Примечание. • р < 0,05; ** р < 0,01; ***р < 0,001.
Следовательно, положение о наличии взаимосвязи между чувствительностью и силой нервной системы находит себе подтверждение не только при применении адекватных раздражителей, но и в том случае, когда в качестве показателя чувствительности нервных клеток выступает электровозбудитель зрительного индикатора.
Рассмотрим теперь результаты сопоставления с силой нервной системы показателей динамики кривых КЧФ как функции величины стимуляции. Все индивидуальные кривые изменения критической частоты мелькающего фосфена в зависимости от амплитуды импульса показаны на рис. 50. По данным угашения с подкреплением все испытуемые могли быть распределены по двум основным группам – «сильных» и «слабых», а кроме того, двое испытуемых с промежуточными данными составили группу «промежуточных».
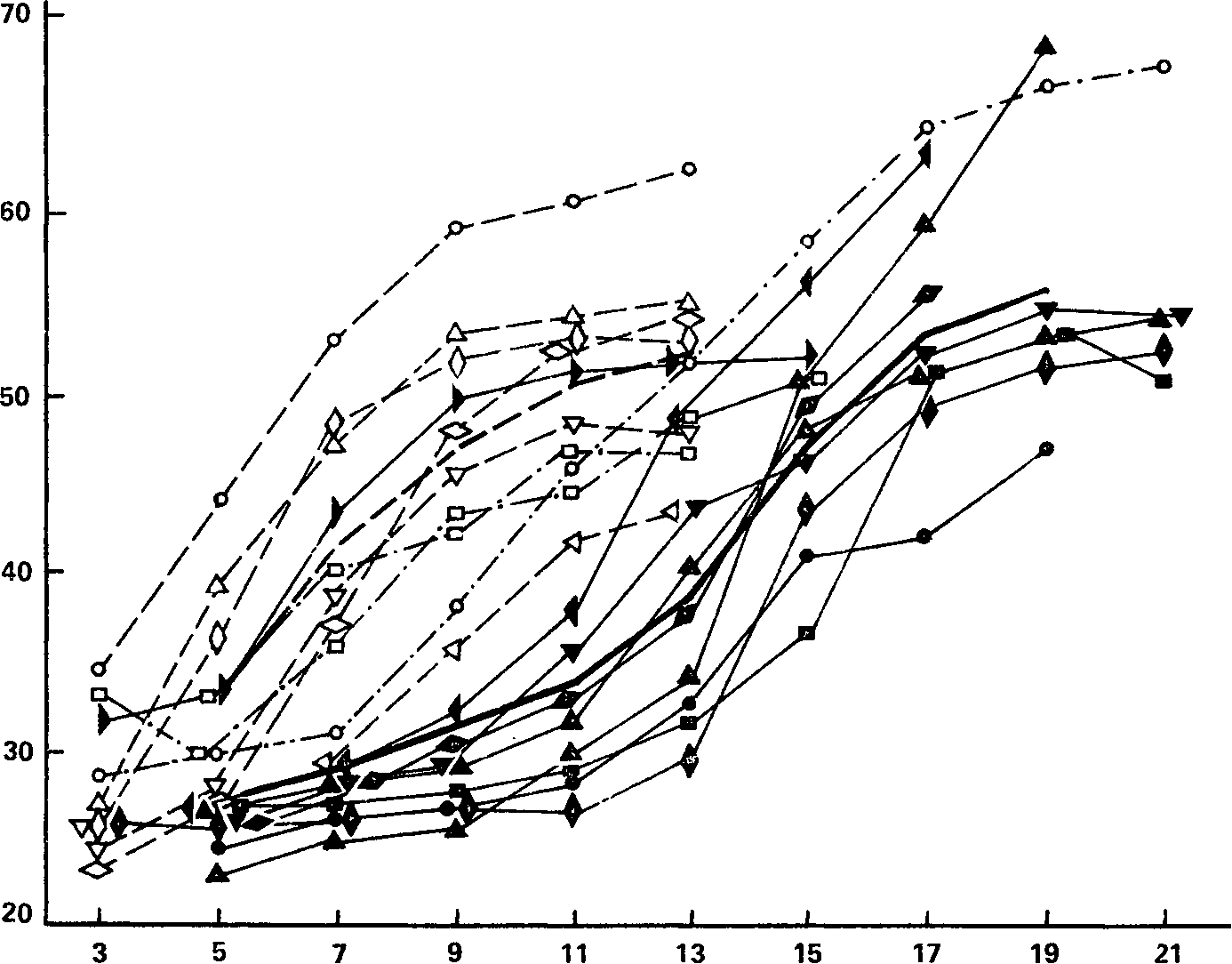
Рис. 50. Критическая частота мелькающего фосфена (КЧФ) как функция интенсивности раздражающего электрического тока. Сплошные. линии принадлежат испытуемым с сильной нервной системой; штриховые линии – испытуемым со слабой нервной системой; штрихпунктирные линии – испытуемым с промежуточным уровнем силы нервной системы. Жирные сплошная и штриховая линии – соответствующие средние.
Ось абсцисс – напряжение (в В); ось ординат – значения КЧФ (В.Д. Небылицын, 1960в).
Кривые испытуемых с сильной нервной системой на рис. 50 обозначены сплошными линиями, со слабой – штриховой, «промежуточных» – штрихпунктиром. Каждую из этих кривых можно характеризовать двумя показателями: во-первых, пределом частоты, т. е. той максимальной высотой, которой достигает кривая при постепенном увеличении интенсивности тока и за которой она у большинства испытуемых практически становится горизонтальной, и, во-вторых, пределом интенсивности, т. е. той величиной раздражителя, при которой достигается частотный предел кривой данного испытуемого. Различия по первому показателю – предельной величине усвоенного ритма – мы пока оставим в стороне, поскольку они, по-видимому, не зависят от силы нервной системы: высота кривой в момент приобретения ею пологости (которая, впрочем, достигнута не у всех испытуемых) может быть самой разнообразной у испытуемых как с сильной, так и со слабой нервной системой. Что же касается различий по второму показателю, то они, видимо, прямым образом связаны с силой нервной системы, и это видно уже из непосредственного сравнения «сильных» и «слабых» кривых.
В самом деле, сплошные кривые, принадлежащие испытуемым с сильной нервной системой, имеют перегиб35, демонстрируя сначала положительное, а затем отрицательное ускорение, и приближаются (кроме одной) к пределу лишь при интенсивности 17 – 21 В. Некоторые кривые «слабых» испытуемых также показывают кратковременное положительное ускорение, но оно заканчивается уже в интервале 3 – 7 В, а при дальнейшем усилении тока эти кривые (как и другие «слабые» кривые) быстро набирают высоту и приобретают пологость уже при напряжениях 11 – 13 В, т. е. уже в тот момент, когд^ кривые «сильных» индивидов только еще начинают свой перегиб. Таким образом, в диапазоне 5 – 13 В все кривые испытуемых со слабой нервной системой имеют отрицательное ускорение и большую высоту над осью абсцисс, а кривые всех испытуемых с сильной нервной системой, за исключением одного, имеют положительное ускорение и малую высоту над абсциссой. В чем причина этой противоположности вида кривых в данном диапазоне стимуляции?
Логично было бы предположить, что причина здесь заключается в различной физиологической действенности физически равных раздражителей, падающих на различные по своей чувствительности сильные и слабые нервные системы (вспомним, что это последнее различие выше было подтверждено статистически).
У более чувствительных слабых нервных систем один и тот же раздражитель (в нашем материале ток напряжением 9 – 13 В) вызывает эффект, близкий к предельному, у менее чувствительных сильных нервных систем – эффект, от предельного еще весьма далекий. Общим результатом этой разницы и является то, что в одном и том же диапазоне раздражителей (5 – 13 В) «сильные» и «слабые» кривые изменения КЧФ имеют существенно различную высоту над осью абсцисс.
Эти различия весьма хорошо коррелируют с силой нервной системы. В табл. 38 представлены коэффициенты корреляции рангов между частотой исчезновения мелькающего фосфена при четырех интенсивностях раздражителя (КЧФ 7, 9, 11, 13 В) и результатами определения силы нервной системы по четырем применявшимся методикам. Из таблицы видно, что эти корреляции достаточно высоки.
Возвращаясь к предположениям, высказанным в начале этого раздела относительно роли силы нервной системы, а точнее, связанных с нею характеристик чувствительности и – гипотетически – лабильности в динамике кривых КЧФ, следует, видимо, признать, что индивидуальные вариации функции КЧФ при изменении интенсивности стимуляции идут именно в том направлении, которое диктуется взаимосвязью между силой и абсолютной чувствительностью. Если бы это было не так, если бы основную роль играла предполагаемая связь между силой и лабильностью, то тогда, очевидно, распределение кривых «сильных» и «слабых» испытуемых на рис. 50 было бы существенно иным.
Однако влияние фактора лабильности нервной ткани на характер кривых КЧФ все же существует. По-видимому, продуктом именно этого влияния являются уже упомянутые выше индивидуальные различия в частотном пределе кривых, явно не зависящие от силовой характеристики индивида. Судя по кривым, максимально достигаемая испытуемыми частота исчезновения фосфена колеблется в среднем около 50 – 55 имп/с, что хорошо совпадает с данными других авторов (Е.Н. Семеновская, 1963). Можно предполагать, что лабильность элементов зрительного анализатора, насколько она может быть измерена приемами определения КЧФ, является независимым по отношению к силе нервной системы фактором мозговой деятельности. К этому вопросу позже мы еще вернемся.
Таблица 38
Коэффициенты корреляции рангов между силой нервной системы, определенной по четырем методикам, и критической частотой мелькающего фосфена на четырех ступенях интенсивности раздражителя (В.Д. Небылицын, 1960в)
Методики определения силы |
КЧФ, В |
|||
7 |
9 |
и |
13 |
|
Угашение с подкреплением |
81*** |
80*** |
84*** |
74*** |
«Утомление» |
83*** |
81*** |
82*** |
79*** |
«Повторение» |
68** |
69** |
71** |
57* |
«Форма кривой» |
63** |
60** |
61** |
52* |
Примечание. • p < 0,05; ** р < 0,01; ***p < 0,001. 253
Новое подтверждение того, что кривая КЧФ при вариациях интенсивности импульса есть показатель силы нервной системы, было получено З. Г. Туровской (19636) в работе по сравнительному изучению ряда индикаторов некоторых свойств нервной системы. Показатель наклона кривой КЧФ коррелировал в этой работе с двумя вариантами индукционной методики и с другими силовыми индикаторами; почти все коэффициенты были статистически значимыми, два из них – на 1 %-ном уровне. Однако порог фосфена в этой работе фактически не коррелировал ни с одним показателем силы нервной системы. Последнее обстоятельство можно объяснить тем, что измерение порога фосфена в экспериментах З.Г. Туровской проводилось лишь в двух-трех опытах, а этого для определения такой нестабильной характеристики, видимо, недостаточно (напомним, что в наших опытах значение порога фосфена выводилось как среднее из 4 опытов, шедших уже после значительной тренировки).
Подводя итоги сказанному относительно роли силы нервной системы в функции КЧФ при измерении интенсивности стимула, нетрудно заметить, что полученные здесь результаты в значительной степени аналогичны результатам эксперимента с временем реакции и так же, как последние, видимо, вполне удовлетворяют тем следствиям из правила взаимосвязи между чувствительностью и силой, которые были сформулированы в начале этой главы. Действительно, при малых – близких к среднему порогу – интенсивностях раздражения у испытуемых с сильной нервной системой наблюдается значительно меньший реактивный эффект, чем у «слабых» индивидов, затем – при возрастании стимула – «слабые» опережают «сильных» в движении функции к пределу и раньше достигают его, и лишь затем при значительно больших величинах стимула к пределу приближаются кривые индивидов с сильной нервной системой. Все эти различия обусловлены различиями самих нервных систем по уровню абсолютных порогов.
Нужно сказать, однако, что в одной из работ, специально проведенной с целью сопоставления методики КЧФ с другими индикаторами силы нервной системы, эта методика, как оказалось, не коррелировала ни с одним из этих индикаторов (В.Д. Небылицын и др., 1965). В объяснение этого факта были выдвинуты несколько предположений, из которых, пожалуй, самое основательное заключается в том, что все силовые индикаторы в этой работе относились к слуховому анализатору, а методика КЧФ, естественно, является индикатором силы нервных клеток зрительного анализатора. Кроме того, могло сыграть свою отрицательную роль и то обстоятельство, что кривые КЧФ получались в данной работе в одном лишь опыте, без всякой предварительной тренировки. Наконец, не исключено, что в данной выборке испытуемых могла проявиться та тенденция «сильных» испытуемых к более высокой лабильности (а «слабых» – к более низкой), о которой говорилось в начале этого раздела и которой достаточно обозначиться лишь у некоторых испытуемых, чтобы свести к нулю обычно наблюдаемую корреляцию.
Таким образом, исследование показателей электровозбудимости глаза в аспекте проблемы, обсуждаемой в настоящей главе, дает подчас недостаточно определенные результаты. Это, очевидно, во многом связано со спецификой самих исследуемых параметров этой психофизиологической функции, которые, с одной стороны, могут быть определены только в результате оценки испытуемым своих иногда трудноуловимых ощущений и в которых, с другой стороны, действуют, вероятно, влияние целой группы специфических для этой методики факторов.
Последнее, надо полагать, справедливо и для той физиологической функции, которая известна под названием навязывания ритма и которая, наряду со временем реакции и критической частотой мелькающего фосфена, была использована нами для проверки следствий, вытекающих из положения о связи между силой нервной системы и абсолютной чувствительностью.