

Внутренние структуры прокариотическофй клетки
 Содержимое
клеток прокариот ограничено от окружающей
среды цито-плазматической мембраной.
Цитоплазматическая
мембрана
– это поли-функциональная органелла,
которая обеспечивает структурную
целостность клетки прокариот и является
площадкой для иммобилизации ферментов,
а также транспортных, регуляторных и
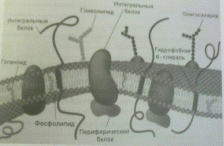
других белков. Соответствует – жид-костно
мозаичной модели (два зеркально
ориентированных слоя фосфолипи-дов и
белков ( периферические, интегральные,
поверхностные) унитарной мембраны у
бактерий и большинства архей. На долю
цитоплазматической мембраны приходится
8-15% сухого вещества клеток. Она состоит
из воды (20%), 80% занимают другие вещества
белки ( 50-60%), липиды (30-50%), полисахариды
(20%), тейхоевые кислоты и неорганические
ионы.
Содержимое
клеток прокариот ограничено от окружающей
среды цито-плазматической мембраной.
Цитоплазматическая
мембрана
– это поли-функциональная органелла,
которая обеспечивает структурную
целостность клетки прокариот и является
площадкой для иммобилизации ферментов,
а также транспортных, регуляторных и
других белков. Соответствует – жид-костно
мозаичной модели (два зеркально
ориентированных слоя фосфолипи-дов и
белков ( периферические, интегральные,
поверхностные) унитарной мембраны у
бактерий и большинства архей. На долю
цитоплазматической мембраны приходится
8-15% сухого вещества клеток. Она состоит
из воды (20%), 80% занимают другие вещества
белки ( 50-60%), липиды (30-50%), полисахариды
(20%), тейхоевые кислоты и неорганические
ионы.
Строение цитоплазматической мембраны бактерий ( по учебнику «Микробиология в определениях и иллюстрациях»)
Цитоплазматическая мембрана выполняет различные функции:
- поддерживает осмотический баланс клетки;
- осуществляет различные виды транспорта;
- участвует в делении клетки;
- биосинтезе внеклеточных полимеров, репликация и сегрегация дочерних хромосом, в векторном транспорте ДНК в клетку;
- в процессах трансформации и конъюгации;
- способствует проникновению вирусов в клетку;
- ассимилирует энергию в форме Pmf и Smf;
- обеспечивает коммуникативные процессы и т.д.
Фосфолипиды в билипидном слое – производные оптически активного D- изомера 3-фосфоглицерола, содержащие два ( редко один) остатка жирных кислот, состав которых весьма разнообразен. В основном это неразветв-ленные мононенасыщенные или жирные насыщенные кислоты с 16-18 атомами углерода в молекуле. Остатка жирных кислот образуют гидрофоб-ную часть молекулы фосфолипида, известную как «гидрофобный хвост». В свою очередь «полярная голова» фосфолипида формируется в результате присоединения к остатку фосфорной кислоты 3-фосфоглицерола или азотного остатка ( этанолами, серии, ходи и др) или остатка полиспиртов (инозитол и др).
Липиды у архей особого типа. Они представляют собой простые эфиры, которые образуются конденсацией глицерола с полиизопреноидными спир-тами с С20 или С23 атомами углерода в молекуле и называются археолами. У архей полярные липиды могут быть представлены простыми тетраэфирами – кальдаархеолами. Они состоят из двух молекул глицероза, связанных с двумя молекулами С 40 – бифитанильных цепей, образованных путем ковалентного соединения «голова к голове» боковых цепей двух диэфиров. В каждую из двух С40 – биитанильных цепей могут быть встроены кольца в количестве от 1 до 4 на углеродную цепочку.
Интерпротоплазматические мембраны – специализированные эндомем-бранные агрегаты, состоящие из поторяющихся модулей сферической, труб-чатой, пластинчатой или дисковидной формы. Основной функцией таких мембран является ассимиляция энергии в форме Pmf. В редких случаях они принимают участие в конструктивном метаболизме. Присутствует посто-янно у цианобактерий, а также характерны для пурпурных бактерий, хемолитотрофов и других.
Цитоплазма - содержимое клетки,которое ограничено от окружеющей среды цитоплазматической мембраной. Представляет собой коллоидную систему. В нее входит цитозоль ( гомогенная система, 70-80 % которой составляет вода, также имеется растворенная РНК), растворенные ферменты, углеводы, жирные кислоты, продукты и субстраты метаболизма. Другая часть представляет собой структурные элементы – органеллы, запасные вещества микрокристаллы, рефрактивные теле и др.
Клетки прокариот не имеют ядра. Но у них имеется образованное равное хромосоме, основу которой составляет компактно упакованная ДНК и называется нуклеоидом. Хромосома прокариот локализована в цитоплазме и не отделена от нее цитоплазматической мембраной. В компактной упаковке ДНК участвуют молекулы РНК, стягивающей петлей; фермент гираза и специфические белки, играющие роль «каркаса». Главную роль в компактизации ДНК играют топоизомеразы, которые образут петли, размыкают и переплетают кольца ДНК. ДНК у прокариот в развернутом виде составляет 0,05-3мм.
Хромосомы являются носителем генома прокариот (совокупность генов, характерная для клеток прокариот. Носителями являются хромосомы. Самый маленький геном у 159 т.п.н у Candidatas Carconella ruddei, большой 9,5 млн п.н у Myxcoccus Xanthus. Если имеется одна хромосома, называется унитар-ным геномом.)
Хромосомы расположены в зоне, где сосредоточены процессы реплика-ции ( это процесс удвоения ДНК, в котором цепи родительской ДНК, содержащие одинаковую информацию, раскручиваются и разделяются с образованием репликативной вилки), репарация ( это полное или частичное восстановление исходной структуры молекулы ДНК, поврежденной при воздействии каких-либо физических или химических факторов), транс-крипции ( это процесс синтеза иРНК с использованием ДНК в качестве матрицы), трансляции ( это синтез полипептидной цепи из аминокислот на рибосоме в соответствии с генетической информацией, записанной в виде последовательности нуклеотидов в молекуле и РНК.
Осуществляется при участии тРНК и связанные с ними метаболиты ( это конечные или промежуточные продукты ( интермедикаты ), образующиеся в клетке в процессе метаболизма).
Хромосома присутствует в клетке в кольцевой, реже линейной форме. В одной клетке прокариот может быть одна или две хромосомы, которые исидентичных групп сцепления.
У представителей фила Planciomyceles, рода Planetomyces, рода Gemmata и рода Isosphaera имеется ядерное тело. Это называется квази-эукарио-тизмом-ограничение ДНК в клетках некоторых прокариот одинарной или двойной цитоплазматической мембраной с образованием структуры , называемой ядерным телом.
Рибосомы - в клетках прокариот представляют собой мультимолеку-лярные комплексы, расположенные в цитоплазме клетки и выполняющие роль универсального аппарата для синтеза белка из аминокислот по заданной матрице на основе генетической информации, продоставляемой иРНК, (трансляция). Рибосомы состоят из рРик, играющих роль в организации рибосомы и рибосомных белков, в массовом соотношении 2:1. Прокариотная рибосома построена из двух субъединиц: большой L-, и малой рРик. Каждая субъединица рибосомы представляет собой свернутый рибонуклеопротеид-ный тяж.
Дивисома – система микрофиламентов у прокариот, которая формируется в сайте деления клетки на основе кольца деления и состоит из белков нес-кольких типов. Дивисома обеспечивает цитокенез – обособление сестринс-ких хромосом внутри своих цитоплазматических компартментов.
Клетки прокариот характерезуется наличием эндоскелета и экзоскелета. Эндоскелет - это система белковых фибрилл, локализованных под цитоплаз-матической мембраной и отвечающих за форму клеток прокариот. Например, антиноподобные белки MreB или Mb1 образуют эндоскелет, который отве-чает за палочковидную форму клетки, а цитоскелетный белок CreS обеспе-чивает вибриоидную или спиральную форму бактерии Caulobacter crescentus. А экзоскелет представлен разнообразными ригидными клеточными струк-турами, расположеными в переплазматическом пространстве и/или за пределами цитоплазматической мембраны. Имеют разный химический состав. У бактерий грамотрицательным морфотипом экзоскелет находится в периплазматическом пространстве и представлен муреиновыми или белко-выми саккулусом (филы Chlamydiae b Thermomierobia). Бактерии с грамполо-жительным морфотипом образуют экзоскелет в виде муреин - тейхоевого саккулуса или муреин-миколил арабииогалактанового саккулуса (фила Actinobacteria) или паракристаллического поверхностного слоя (S-слой). У архей экзоскелет представлен белковым S-слоем, псевломуреиновым саккулусом или гетерополисахаридным слоем. У членов филы Spirochaetes экзоскелетом является осевая структура, образованная пучком периплазма-тических жгутиков. Корбоксисомы - имеются в клетках автотрофных бактерий ( это организмы, обладающие системой взаимосвязанных анаболических реакций. Обеспе-чивающих усвоение углекислого газа и ряда других одноуглеродных субстратов в качестве единственного или главного источника углерода). Представлены многогранными структурами. Окруженными белковой мембраной. Содержат плотно упакованные молекулы фермента-рибулозобифосфат-карбоксилазу (РуБисКо), катализирующего ассимиляцию углекислого газа в цикле Кальвина – Бенсона. Карбоксисома содержит до 300 молекл РуБисКо. Упакованных в виде паракристаллической структуры. Сложноорганизованные, инертные структуры прокариот, окруженные однослойной белковой неунитарной мембраной и заполненные газом, аналогичным по составу газу в окружающей среде, называются газовыми визикулами. Обеспечиват водным прокариотам пассивную подвижность за счет изменения плавучести и придают им способность оставаться в воде во взвешанном состоянии.
Тиллакоиды - это интрацитоплазматические мембраны которые имеют фотосинтетический аппарат (полипептидный комплекс, преобразует погла-щенную энергию квантов света в Pmf способны к фототрофии) у циано-бактерий (одноклеточные, колониальные бактерии с услажненным грамо-трицательным морфотипом. Облегатные или факультативные фототрофы). У фототрофных бактерий (аноксигенные фототрофы) имеют также хлоросомы- это светособирающие перефирические антены ,в виде пузырька на внутренней поверхности цитоплазматической мембраны, окруженная неунитарной мембраной. Пигменты, локализованные в хлоросоме, погло-щают кванты света и передают их энергию в реакционные центры. Протеосомы - это мультиферментный комплекс, служащий для уничтожения путем протеолиза белков (ферментативное ращепление белков при участии протеаз-протеолитические ферменты, относящиеся к классу гидролаз, ката-лизирующие гидролиз белков и пептидов, с разрывом пептидных связей между определенными аминокислотами), которые под воздействием срессовых факторов или по возрасту утратили свои функциональные свойства.
Экзосома имеется у архей. Выполняют функцию утилизации РНК. У планктоминетов (не у всех) в клетке имеется анаммоксосома – это дыхатель-ная органелла, окруженная унитарной мембраной, Построенной из плотно упакованных липидов-леддеранов. Она запасает энергию для клетки за счет анаэробного окисления аммиака с использованием в качестве конечного акцептора электронов нитрит – анион. Анаммоксосома занимает до 30 % внутреннего объема клетки. В мембране присутствует АТФ-синтеза. Ведущая фосфорилирование АДФ при движении протонов по градиенту их концентрации, т.е. из анаммоксосомы в цитоплазму. Магнетосама – ограниченная унитарной мембраной органелла бактерий, заполненная электронно-плотными кристалами соединений железа в виде магнетита, или грейтита, или пирротита, или пирита. Кристаллы имеют форму: усеченных октаэдров, гексагональных и прямоугольных призм и тому подобное. Обеспечивают магнитную ориентацию клетки в водных экосистемах и избыточно увлажненных верхних горизонтах почв. Это позволяет бактериям покидать неблагоприятные для жизнедеятельности зоны и находить места с благоприятными микроаэрофильными условиями и необходимыми для роста органическими субстратами.
В определенных условиях прокариоты накапливают запасные вещества в осматически инертной форме (не растворимые в воде). Они представляют собой динамическое депо, в котором прокариотная клетка запасает или энергию или конструктивный субстрат или восстановитель.
К запасным вещам относят: 1. Серные глобулы - это включения округлой формы в клетках прокариот, ограниченные белковой неунитарной мембраной и содержащие серу, либо в форме циклооктосеры,либо политионата или серных цепей. Откладываются в цитоплазме или периплапзматическом пространстве в качестве промежу-точного продукта при окислении экзогенного донора электронов –сульфида. Сера используется бактериями в качестве эндогенного донора электронов при истощении запасов сульфида в местах обитания. 2. Цианофициновые гранулы - это сферические структуры, состоящие из белка в состав которого входит аспаргиновая кислота и L-аргинин в эквимолярных количествах. Характерны только для некоторых цианобак-терий. В темноте используется как резервный источник энергии, а на свету - в условиях стресса служат источноком азота.
3.Гликоген - это разветвленный полисахарид, молекулы которого построены из остатков а – D- глюкозы. Это быстро мобилизируемый энергетический резерв бактерий. При гидролизе гликоген ращепляется через ряд промежуточных продуктов до глюкозы.
4. Крахмал представлен полигликозидными твердофазными гранулами, состоящими из двух глюканов: амилозы и амилопектина. Является резервным источником углерода и энергии у некоторых прокариот. 5. Гранулоза представлена полигликозидными мелкими гранулами, построенными из остатков а-D-глюкозы. Откладывается в клетках у Clostredium перед началом образования эндоспор в количестве до 60% сухого вещества. Резервный источник углерода и энергии. 6. Трегалоза – это дисахарид, построенный из двух молекул глюкозы , соединенных а-1, 1-гликозидной связью. Откладывается как запасное вещество.
В цитоплазме некоторых членов домена Bacteria присутствуют спирали-зованные белковые ламеллы, которые называются рефрактильными телами или R-телами. Являются симбионтами простейших, всмешанной популяции простейших , которая состоит из незаряженных особей и носителей бакте-риальных симбионтов, бактерии делают своих хозяев эффективными убий-цами, что дает преимущество в борьбе среди сородечей. Токсины, продуци-руемые бактериями. Вызывают либо лизис, либо параличчуствительных особей простейших. Эндотоксины – это интегральные компоненты клеточной стенки бактерий с грамотрицатльным морфотипом, освобождаю-щиеся большей частью после гибели микроорганизмов. По химической природе являются комплексом белков, липидных и полисахаридных остатков. Токсигенные свойства эндотаксина определяются его целой молекулой. Она обладает стабильностью, термостабильны, менее токсичны и имеют иммуногенность. При поступлении в организм в большинстве угнетают активность фагацитов, гранулоцитов, моноцитов, увеличивают проницаемость капилляров, оказывают разрушительное действие на клетки. Эндотоксины стимулирует синтез интерферонов, активируют систему комплимента по классическому пути. Обладают свойствами аллергенов. При поведении эндотоксина в небольших дозах повышается резистентность организмам. К эндотоксинам относят токсины возбудителей : брюшного тифа, бруцеллеза и др.
Кристаллические включения. Некоторые протобактерии проводят детоксикацию поступающих в клетку ядовитых соединений – селена, теллура. И преобразуют в балластные включения в цитоплазме. К ним относят: Микрокристалы кальцита - специфические цитоплазматические включения, не ограниченные мембраной и занимающие до 70% объема клетки бактерии Achromatium oxaliferum.
Функциями являются: нейтрализация кислотности, возникающий при окислении серы; повышение внутриклеточного парциального давления углекислого газа для облегчения автотрофного роста; увеличение интенсив-ности транспортных процессов. Микрокристаллы селена – специфические цитоплазматические балластные включения элементного селена в виде округлых кристаллов; не ограничен-ных мембраной. Образуются некоторыми преставителями бактериальных фил – Proteobacteria и Firmicutes и археотных фил Crenarehaeota и Euryarchaeota с целью детоксикации поступающего в клетку из внешней среды селенита. Микрокристаллы теллура – специфические цитоплазматические балластные игольчатые кристаллы элементного теллура, не ограниченные мембраной. Присутствуют в клетках квази-фототрофных бактерий из филы Proteobacteria, родов Erythrobacter, Erythromicrobium, Roseococcus др. образуются в результате восстановления ядовитого аниона теллурита до элементарного теллура с целью сохранения клетки в жизнеспособном состоянии.
Эндоспора - сложные специализированные покоящееся формы бактерий. Формируются эндогенно внутри цитоплазмы материнской клетки путем – «поглащения» одной клетки (будущей эндоспоры), другой ( материнской). Для них характерны: специфические многослойные покровы, наружая и внутренняя унитарные мембраны, кортекс, высокая устойчивость к действию разнообразных абиотических факторов.