

Источники генерации ээг
Временные параметры ритмических составляющих ЭЭГ исчисляются мс×10, мс×102. Поскольку длительность ПД нейрона составляет не более 2 мс и не сопоставима с ЭЭГ по временным параметрам, считается, что импульсная активность нейронов не находит отражения в колебаниях потенциала, регистрируемого с поверхности мозга или со скальпа.
Принято считать, что в ЭЭГ находит отражение синаптическая активность нейронов. ВПСП и ТПСП имеют значительную длительность (>30 мс и >70 мс соответственно), не подчиняются закону "все или ничего", имеют градуальный характер и могут суммироваться. Упрощая картину, можно сказать, что колебания+ потенциала на поверхности коры связаны с ВПСП в ее глубинных слоях либо с ТПСП в поверхностных слоях, а колебания– предположительно отражают противоположное соотношение источников электрической активности.
Ритмы
Ритмический характер биоэлектрической активности коры (в частности α-ритма) обусловлен в основном влиянием таламуса, где локализованы главные, но не единственные пейсмекеры (нейроны неспецифического таламуса). Одностороннее удаление или хирургическая изоляция таламуса от неокортекса приводит к полному исчезновению α-ритма в ипсилатеральных зонах коры, тогда как ритмическая активность самого таламуса не меняется. Ретикулярная формация ствола мозга может оказывать синхронизирующее и дезинхронизирующее влияние, т.е.
В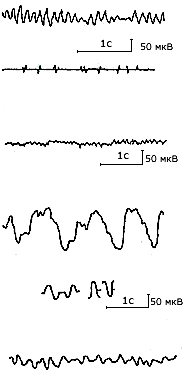 ыделяют
следующие ритмы мозга.
ыделяют
следующие ритмы мозга.
Альфа-ритм (8-13 Гц, 5-100 мкВ) регистрируется преимущественно в затылочной и теменной областях, обычно модулирован в веретена. Проявляется при закрытых глазах в состоянии покоя, медитации или при монотонной деятельности. Предполагается, что альфа-ритм отражает реверберацию возбуждений, кодирующих внутримозговую информацию и создающих оптимальный фон для процесса приема и переработки афферентных сигналов. Его роль состоит в своеобразной функциональной стабилизации состояний мозга и обеспечении готовности реагирования. Предполагается также, что альфа-ритм связан с действием селектирующих механизмов мозга, выполняющих функцию резонансного фильтра, и таким образом регулирующих поток сенсорных импульсов.
Бета-ритм (18-30 Гц, 2-20 мкВ) регистрируется в прецентральной и фронтальной коре. Сопровождает переход от состояния покоя к напряжению, причем значимое усиление высокочастотной активности наблюдается при умственной деятельности, включающей элементы новизны. Связан с максимальным сосредоточением внимания при решении сложных задач.
Гамма-колебания (от 30 до 120-170 Гц или до 500 Гц, 2 мкВ) регистрируется в прецентральной, фронтальной, височной, теменной и специфических зонах коры.
Дельта-волны (0,5-4,0 Гц, 20-200 мкВ), возникает в разных участках коры мозга при глубоком естественном и наркотическом сне (четвертая стадия сна) и может усиливаться при выполнении мыслительных заданий (!).
Тета-волны (4-7 Гц, 5-100 мкВ) наблюдаются во фронтальных зонах, в гиппокампе, чаще двухсторонние синхронные, иногда (особенно в лобной области) с веретенообразной модуляцией. При открытых глазах и умственной нагрузке распространяется и на другие зоны коры. Тэта-ритм тесно связан с эмоциональным и умственным напряжением (стресс-ритм) и сопровождает переживание как +, так и - эмоций. Многие ученые связывают его наличие с поисковым поведением, с извлечением энграмм из памяти и с планированием деятельности, причем усиление этой составляющей положительно соотносится с успешностью решения задач. По своему происхождению тэта-ритм связан с кортико-лимбическим взаимодействием. Предполагается, что усиление тэта-ритма при эмоциях отражает активацию коры больших полушарий со стороны лимбической системы.
Лямбда-колебания (12-14 Гц, 20-50 мкВ или 100-200 мкВ на максимуме), их фокус приходится на вертекс. Регистрируется при переходе дремоты в медленноволновыи сон, а также при раздражении неспецифических ядер таламуса.
Сигма-активность (сонные веретена) (12-14 Гц) имеют широкую зону распространения.
Мю-ритм (роландический, или аркообразный), имеет ту же частоту, что и альфа-ритм, но другую локализацию, и чувствителен к другим видам модальности; отвечает блокадой на проприоцептивные раздражения; регистрируется в области роландовой борозды.
Каппа-колебания (8-12 Гц, 5-40 мкВ) наблюдаются в височной области при подавлении альфа-ритма умственной деятельностью.
Тay-ритм регистрируется в височной коре и подавляется звуковыми стимулами.
Кроме того, описаны сверхмедленные потенциалы, имеющие период колебаний от нескольких секунд до нескольких часов и амплитуду от сотен микровольт до десятков милливольт. Эти потенциалы составляют 5 групп: 1 - секундные с периодом колебания от 3 до 10 сек., 2 -декасекундные с периодом 15-60 сек., 3 - минутные с периодом 2-9 мин., 4- декаминутные с периодом 10-20 мин., 5 - часовые с периодом 0,5 -1,5 часа. Подобные колебания имеют место и при бодрствовании, и во время сна, при извлечении энграмм из памяти, при действии фармакологических веществ, а также при патологии мозга.