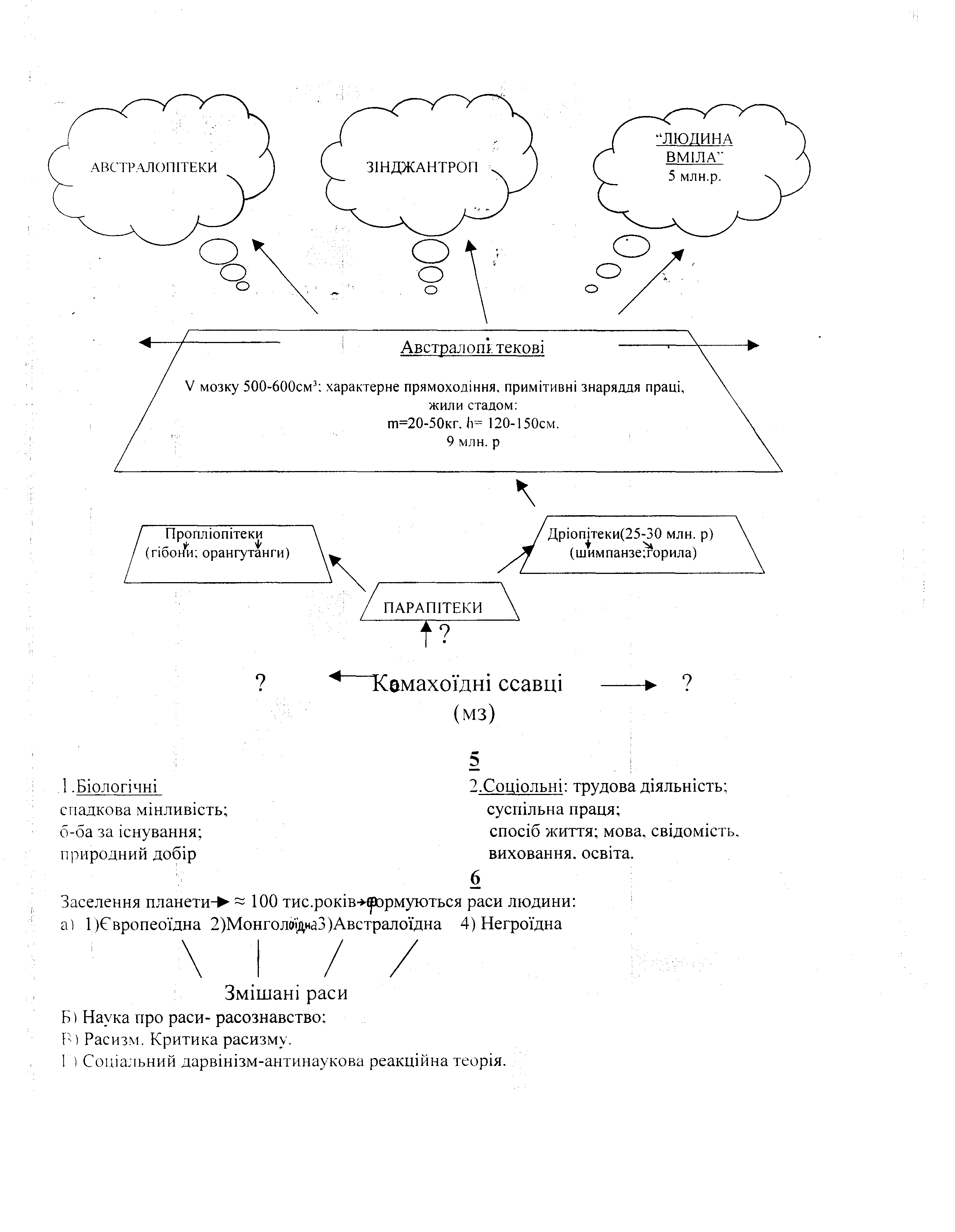
Історія дослідження
Термін «фермент» був запропонований у 17 столітті хіміком ван Гельмонтом для опису механізмів травлення. В кінці 18 — на початку 19 століття вже було відомо, що м'ясо перетравлюється шлунковим соком, а крохмаль перетворюється на цукор під дією слини. Проте механізм цих явищ був ще невідомий[1]. В 19 столітті Луї Пастер, вивчаючи перетворення вуглеводів в етиловий спирт під дією дріжджів, дійшов до висновку, що цей процес (бродіння) каталізується якоюсь «життєвою силою», що знаходиться в дріжджових клітинах.
Понад сто років тому терміни «фермент» і «ензим» відображали різні погляди Луї Пастера з одного боку та Марселена Бертло і Юстуса Лібіха з іншого в теоретичній суперечці про природу спиртового бродіння. Власне «ферментами» (від лат. fermentum — «закваска») називали «організовані ферменти» (тобто саме живі мікроорганізми), а термін «ензим» (від грец. ἐν- — «в-» і ζύμη — «дріжджі», «закваска»), запропонований 1876 року В. Кюне для «неорганізованих ферментів», що секретуються клітинами, наприклад, до шлунку (пепсин) або кишечника (трипсин, амілаза). За два роки по смерті Пастера 1897 року Едуард Бюхнер опублікував роботу «Спиртове бродіння без дріжджових клітин», в якій експериментально показав, що екстракт клітин дріжджів здійснює спиртове бродіння так само, як і незруйновані дріжджові клітини[2] 1907 року за цю роботу він був удостоєний Нобелівської премії.
Функції ферментів
Ферменти є біологічними каталізаторами, вони присутні у всіх живих клітинах і сприяють перетворенню одних речовин (субстратів) на інші (продукти). Ферменти виступають в ролі каталізаторів практично у всіх біохімічних реакціях, що відбуваються в живих організмах — ними каталізується біля 4000 хімічно окремих біореакцій[3]. Ферменти грають найважливішу роль у всіх процесах життєдіяльності, скеровуючи та регулюючи обмін речовин організму.
Подібно до всіх каталізаторів, ферменти прискорюють як пряму, так і зворотну реакцію, знижуючи енергію активації процесу. Хімічна рівновага при цьому не зміщується ні в прямий, ні у зворотний бік. Відмінність ферментів від небілкових каталізаторів полягає у їхній високій специфічності — константа дисоціації деяких субстратів з білком-ферментом може досягати менш ніж 10−10 моль/л.
Ферменти широко використовуються і в народному господарстві — харчовій, текстильній промисловості, у фармакології.
Структура і механізм дії ферментів
Активність ферментів визначається їхньою тривимірною структурою.
Як і всі білки, ферменти синтезуються у вигляді лінійного ланцюжка амінокислот, який згортається певним чином. Кожна послідовність амінокислот згортається особливим чином, і молекула (білкова глобула), що виходить, володіє унікальними властивостями. Кілька білкових ланцюжків можуть об'єднуватися у білковий комплекс. Найбільші рівні структури білків — третинна та четвертинна структури — руйнуються при нагріванні або під дією деяких хімічних речовин.
Щоб каталізувати реакцію, фермент повинен зв'язатися з одним або кількома субстратами. Білковий ланцюжок ферменту згортається таким чином, що на поверхні глобули утворюється щілина або западина, до якої приєднуються молукули субстрату. Ця область називається ділянкою (сайтом) зв'язування субстрата. Зазвичай вона співпадає з активним центром ферменту або знаходиться поблизу від нього. Деякі ферменти містять також ділянки зв'язування кофакторів або іонів металів.
У деяких ферментів присутні також ділянки зв'язування малих молекул, що не беруть безпосередньої участі в реакції і часто, але не обов'язково, є субстратами або продуктами метаболічного шляху, в який входить фермент. Вони зменшують або збільшують активність ферменту, що створює можливість для зворотного зв'язку або регуляції роботи ферменту.
Выводы:
Ферменты характеризуются следующими основными свойствами.
1. Все они представляют собой глобулярные белки.
2. Информация о них, как и о других белках, закодирована в ДНК.
3. Ферменты действуют как катализаторы.
4. Их присутствие не влияет ни на природу, ни на свойства конечного продукта реакции.
5. Ферменты действуют чрезвычайно эффективно, т. е. очень малое количество фермента вызывает превращение больших количеств субстрата. Одна молекула каталазы способна, например, при температуре тела разложить за одну секунду на воду и кислород около 600 тысяч молекул пероксида водорода.
6. Ферменты высокоспецифичны, т. е. один фермент катализирует обычно только одну реакцию. Каталаза, например, катализирует только расщепление пероксида водорода.
7. Катализируемая ферментом реакция обратима.
8. Активность ферментов меняется в зависимости от рН и температуры, а также от концентрации как субстрата, так и самого фермента.
9. Ферменты снижают энергию активации катализируемой реакции .
1
 0.
В молекуле фермента есть активный центр,
который вступает в контакт с субстратом.
Этот активный центр имеет особую
форму.
0.
В молекуле фермента есть активный центр,
который вступает в контакт с субстратом.
Этот активный центр имеет особую
форму.
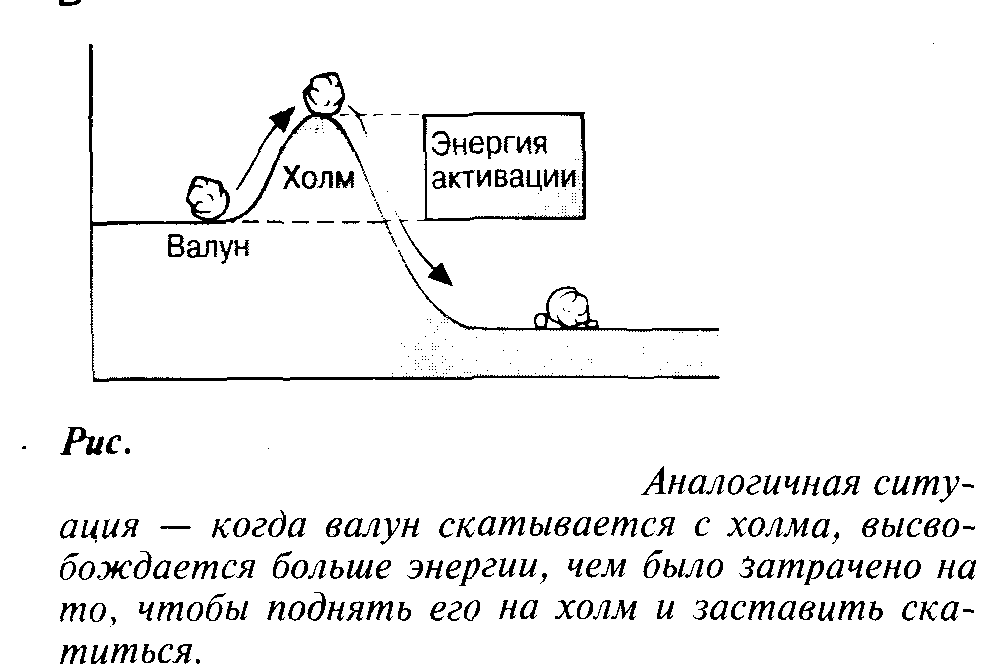
Энергия активации
П
 редставим
себе смесь бензина и кислорода. Реакция
между этими двумя веществами с
термодинамической точки зрения
возможна, но она не пойдет без затраты
некоторого количества энергии,
поступившей, например, в форме простой
искры. Так же обстоит дело и со спичкой.
Содержащиеся в спичечной головке
вещества могут вступить в реакцию,
суммарным итогом которой будет
выделение энергии, но чтобы запустить
реакцию, придется сначала затратить
небольшое количество энергии (достаточно
тепловой энергии, выделяющейся, когда
мы чиркаем спичкой о коробок). Энергия,
необходимая для того, чтобы заставить
вещества вступить в реакцию, называется
энергией
активации. Ферменты,
действуя как катализаторы, снижают
энергию активации (рис.). Они повышают
общую скорость реакции, не изменяя
в сколько-нибудь значительной степени
температуру, при которой эта реакция
протекает.
редставим
себе смесь бензина и кислорода. Реакция
между этими двумя веществами с
термодинамической точки зрения
возможна, но она не пойдет без затраты
некоторого количества энергии,
поступившей, например, в форме простой
искры. Так же обстоит дело и со спичкой.
Содержащиеся в спичечной головке
вещества могут вступить в реакцию,
суммарным итогом которой будет
выделение энергии, но чтобы запустить
реакцию, придется сначала затратить
небольшое количество энергии (достаточно
тепловой энергии, выделяющейся, когда
мы чиркаем спичкой о коробок). Энергия,
необходимая для того, чтобы заставить
вещества вступить в реакцию, называется
энергией
активации. Ферменты,
действуя как катализаторы, снижают
энергию активации (рис.). Они повышают
общую скорость реакции, не изменяя
в сколько-нибудь значительной степени
температуру, при которой эта реакция
протекает.
Механизм действия ферментов
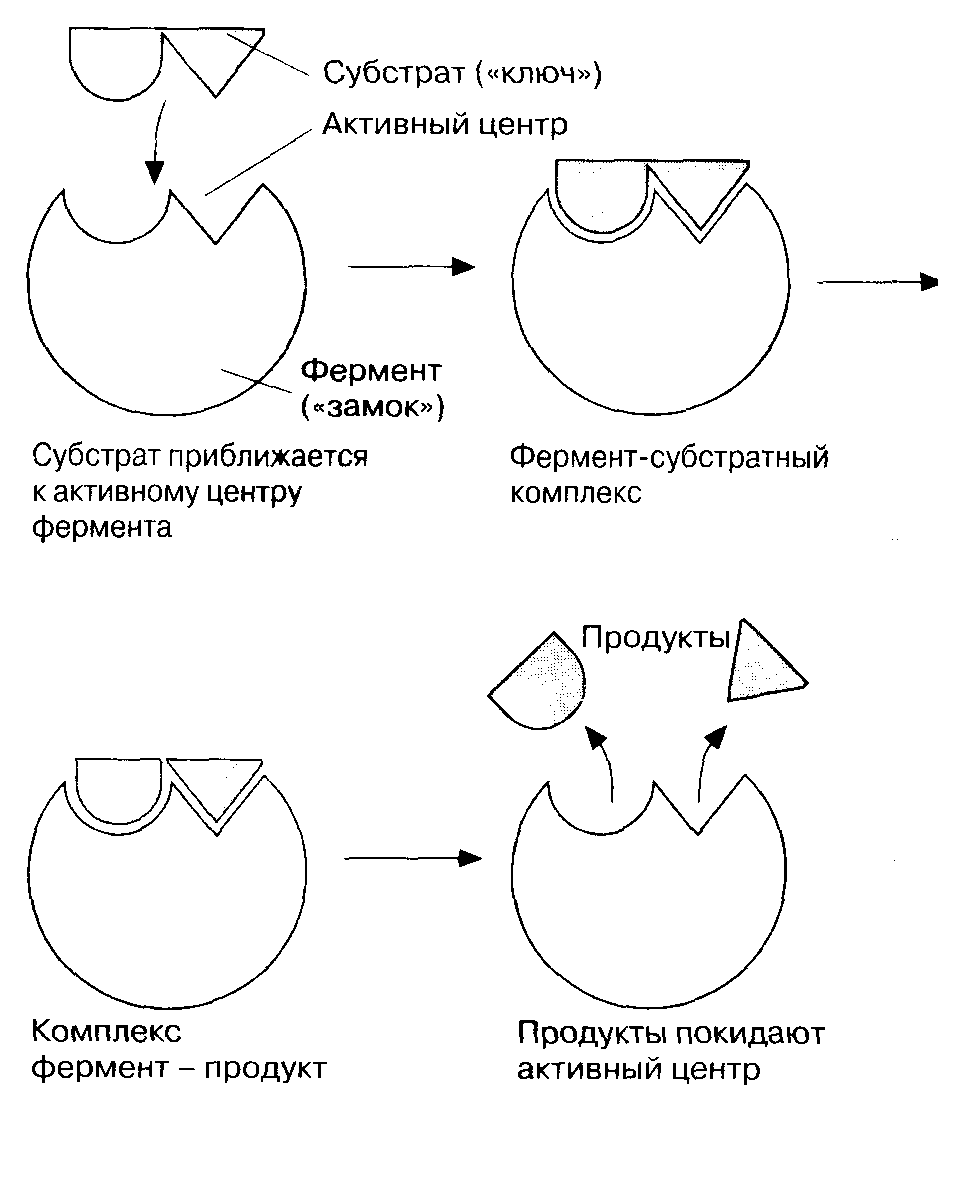
Ферменты обладают очень высокой специфичностью. Фишер (Fischer) в 1890 г. высказал предположение, что эта специфичность обусловливается особой формой молекулы фермента, точно соответствующей форме молекулы субстрата (или субстратов). Эту гипотезу часто называют гипотезой «ключа и замка»: субстрат сравнивается в ней с «ключом», который точно подходит по форме к «замку», т. е. к ферменту. В схематическом виде это представлено на рис. Часть молекулы фермента, вступающую в контакт с субстратом, называют активным центром фермента, и именно активный центр фермента имеет особую форму.
Образовавшиеся продукты по форме уже не соответствуют активному центру фермента. Они отделяются от него (поступают в окружающую среду), после чего освободившийся активный центр может принимать новые молекулы субстрата.
В
 1959 г. Кошланд (Koshland)
предложил новую интерпретацию гипотезы
«ключа и замка», получившую название
гипотезы
«индуцированного соответствия». На
основе данных, позволяющих считать
ферменты и их активные центры физически
более гибкими, чем это казалось вначале,
он заключил, что субстрат, соединяясь
с ферментом, вызывает какие-то изменения
в структуре его активного центра.
Аминокислотные остатки, составляющие
активный центр фермента, принимают
определенную форму, которая дает
возможность ферменту наиболее
эффективным образом выполнять свою
функцию. Подходящей аналогией в этом
случае может служить перчатка, которая
при надевании на руку соответствующим
образом изменяет свою форму.
1959 г. Кошланд (Koshland)
предложил новую интерпретацию гипотезы
«ключа и замка», получившую название
гипотезы
«индуцированного соответствия». На
основе данных, позволяющих считать
ферменты и их активные центры физически
более гибкими, чем это казалось вначале,
он заключил, что субстрат, соединяясь
с ферментом, вызывает какие-то изменения
в структуре его активного центра.
Аминокислотные остатки, составляющие
активный центр фермента, принимают
определенную форму, которая дает
возможность ферменту наиболее
эффективным образом выполнять свою
функцию. Подходящей аналогией в этом
случае может служить перчатка, которая
при надевании на руку соответствующим
образом изменяет свою форму.
Скорость ферментативных реакций
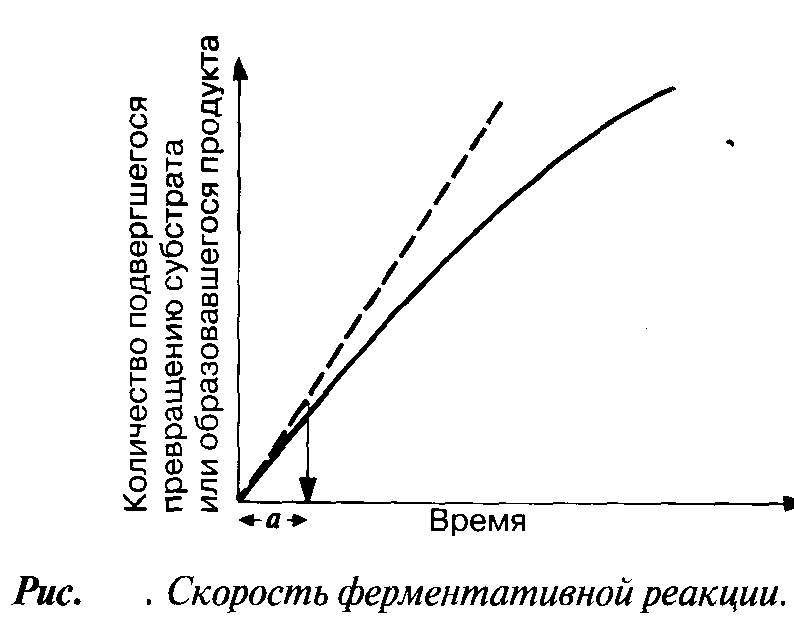
Мерой скорости ферментативных реакций служит количество субстрата, подвергшегося превращению в единицу времени, или количество образовавшегося продукта.
Скорость определяют по углу наклона касательной к кривой на начальной стадии (а на рис.) реакции. Чем круче наклон, тем больше скорость. Со временем скорость реакции обычно снижается, по большей части в результате снижения концентрации субстрата (см. след. разд.).
Концентрация фермента
При высокой концентрации субстрата и при постоянстве других факторов, таких, например, как температура и рН, скорость ферментативной реакции пропорциональна концентрации фермента Катализ осуществляется всегда в условиях, когда концентрация фермента гораздо ниже концентрации субстрата. Поэтому с возрастанием концентрации фермента растет и скорость ферментативной реакции.
Температура
С
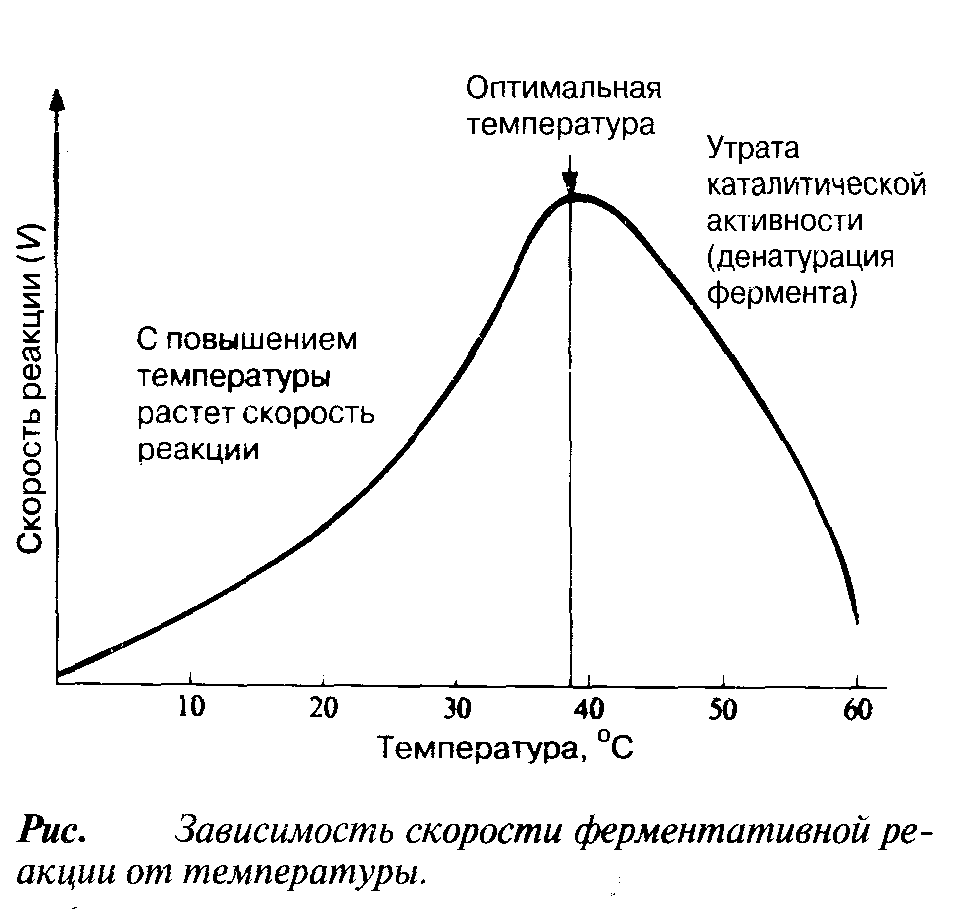 повышением температуры ускоряется
движение молекул, вследствие чего у
молекул субстрата и фермента
оказывается больше шансов столкнуться
друг с другом. В результате увеличивается
и вероятность того, что реакция произойдет.
Температура, обеспечивающая максимальную
активность, называется оптимальной
температурой. Если температура
поднимается выше этого уровня, скорость
ферментативной реакции снижается,
несмотря на увеличение частоты
столкновений. Происходит это вследствие
разрушения вторичной и третичной
структур фермента, иными словами,
вследствие того, что фермент претерпевает
денатурацию (рис.). Молекула фермента
развертывается и его активный центр
постепенно утрачивает присущую ему
форму. Наиболее чувствительны к
воздействию высокой температуры
водородные связи и гидрофобные
взаимодействия.
повышением температуры ускоряется
движение молекул, вследствие чего у
молекул субстрата и фермента
оказывается больше шансов столкнуться
друг с другом. В результате увеличивается
и вероятность того, что реакция произойдет.
Температура, обеспечивающая максимальную
активность, называется оптимальной
температурой. Если температура
поднимается выше этого уровня, скорость
ферментативной реакции снижается,
несмотря на увеличение частоты
столкновений. Происходит это вследствие
разрушения вторичной и третичной
структур фермента, иными словами,
вследствие того, что фермент претерпевает
денатурацию (рис.). Молекула фермента
развертывается и его активный центр
постепенно утрачивает присущую ему
форму. Наиболее чувствительны к
воздействию высокой температуры
водородные связи и гидрофобные
взаимодействия.
Температурный оптимум для большинства ферментов млекопитающих лежит в пределах 37—40 °С. Существуют, однако, ферменты с более высоким температурным оптимумом; у бактерий, живущих в горячих источниках, он может, например, превышать 70оС. Именно такие ферменты используются в качестве добавок к стиральным порошкам для стирки в горячей воде. Когда температура приближается к точке замерзания или оказывается ниже ее, ферменты инактивируются, но денатурации при этом не происходит. С повышением температуры их каталитическая активность вновь восстанавливается.
р
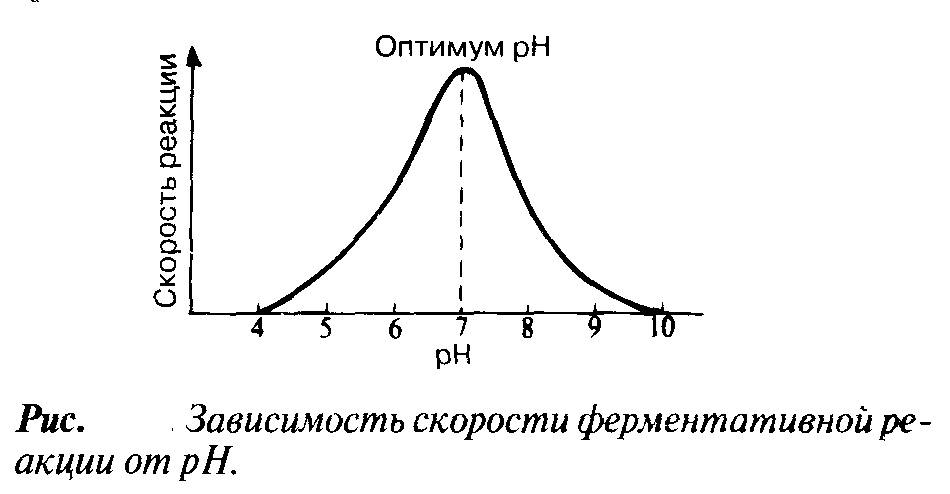 Н
Н
При постоянной температуре любой фермент, как правило, работает наиболее эффективно в узких пределах рН. Оптимальным считается то значение рН, при котором реакция протекает с максимальной скоростью. При более высоких и более низких рН активность фермента снижается. Сдвиг рН меняет заряд ионизированных кислотных и основных групп, что ведет к разрушению ионных связей, участвующих в поддержании специфичной формы молекул фермента. В результате изменяется форма молекул фермента - и в первую очередь форма его активного центра. При слишком резких сдвигах рН фермент денатурирует.
Ингибирование ферментов
Известны различные низкомолекулярные соединения, которые могут снижать скорость ферментативных реакций. Такие соединения называются ингибиторами ферментов. Важно понимать, что ингибирование — это один из нормальных способов регулирования активности ферментов. Многие лекарственные препараты и яды также действуют как ингибиторы ферментов.
Кофакторы ферментов
Многим ферментам для эффективной работы требуются те или иные небелковые компоненты, называемые кофакторами. Кофакторы — это вещества, присутствие которых совершенно необходимо для проявления каталитической активности ферментов, хотя сами они в отличие от ферментов сохраняют стабильность при довольно высоких температурах. Роль кофакторов могут играть различные вещества — от простых неорганических ионов до сложных органических молекул; в одних случаях они остаются неизменными в конце реакции, в других — регенерируют в результате того или иного последующего процесса. Кофакторы подразделяются на три типа: неорганические ионы, простетические группы и коферменты. Их мы и рассмотрим в последующих трех разделах.
Неорганические ионы (активаторы ферментов)
Полагают, что эти ионы заставляют молекулы фермента или субстрата принять форму, способствующую образованию фермент-субстратного комплекса. Тем самым увеличиваются шансы на то, что фермент и субстрат действительно прореагируют друг с другом, а следовательно, возрастает и скорость реакции, катализируемой данным ферментом. Так, например, активность амилазы слюны повышается в присутствии хлорид-ионов.
Простетические группы (например, ФАД, гем)
Если кофактор прочно связан с ферментом и остается в этом связанном состоянии постоянно, то его называют простетической группой. Роль простетических групп играют органические молекулы. Они помогают ферменту осуществлять его каталитическую функцию, как это видно на примере флавинадениндинуклеотида (ФАД). ФАД содержит рибофлавин (витамин В2), который является водород-акцепторной частью его молекулы. Функция ФАД связана с окислительными путями клетки, в частности с процессом дыхания, в котором ФАД играет роль одного из переносчиков в дыхательной цепи.
Гем — это железосодержащая простетическая группа. Его молекула имеет форму плоского кольца (порфириновое кольцо, такое же, как у хлорофилла), в центре которого находится атом железа. Гем выполняет в организме ряд биологически важных функций.
ПЕРЕНОС ЭЛЕКТРОНОВ. В качестве простетической группы цитохромов (см. дыхательную цепь в гл. 9) гем выступает как переносчик электронов. Присоединяя электроны, железо восстанавливается до Fe(II), а отдавая их, окисляется до Fe(III). Гем, следовательно, принимает участие в окислительно-восстановительных реакциях за счет обратимых изменений валентности железа.
ПЕРЕНОС КИСЛОРОДА. Гемоглобин и миоглобин — два гемсодержащих белка, осуществляющих перенос кислорода. Железо находится в них в восстановленной, Fe(II), форме.
КАТАЛИТИЧЕСКАЯ ФУНКЦИЯ. Гем входит в состав каталаз и пероксидаз, катализирующих расщепление пероксида водорода до кислорода и воды. Содержится он также и в некоторых других ферментах.
Коферменты (например, НАД, НАДФ, ацетилкофермент А, АТФ)
Коферменты, как и простетические группы, — это органические молекулы, выполняющие функцию кофакторов, но в отличие от простетических групп они сохраняют связь с ферментом только в ходе реакции. Все коферменты представляют собой производные витаминов.
Никотинамидадениндинуклеотид (НАД) — производное витамина ниацина («никотиновой кислоты») — может существовать как в окисленной, так и в восстановленной форме. В окисленной форме НАД при катализе играет роль акцептора водорода.

Самостійне вивчення 3
1. Генетика людини вивчає явища спадковості і мінливості у популяціях людей, особливості успадкування нормальних і патологічних ознак, залежність захворювання від генетичної схильності і факторів середовища. Генетика людини— галузь генетики, тісно пов'язана з антропологією і медициною. Генетику людини умовно поділяють на антропогенетіку, що вивчає спадковість і мінливість нормальних ознак людського організму, і генетику медичну, яка вивчає його спадкову патологію (хвороби, дефекти, потворність та ін.). Генетика людини пов'язана також з еволюційною теорією, оскільки досліджує конкретні механізми еволюції людини і його місце в природі з психологією, філософією, соціологією. З напрямів генетики людини найінтенсивніше розвиваються цитогенетика, біохімічна генетика, імуногенетика, генетика вищої нервової діяльності, фізіологічна генетика.
2. Уже в 1750 р. французький лікар П.Мопертюї описав характер успадкування багатополості (полідакталії). Проведений ним аналіз успадкування цієї ознакіи багато в чому передував відкриттю Г.Менделя. У 1814 р. Дж.Адамс опублікував працю , в якій розрізняв не спадкові й спадкові захворювання.
За період 1803-1820 р. кілька лікарів описали тип успадкування гемофілії. Швейцарський лікар-офтальмолог Й.Горнер у 1876 р. описав тип успадкування дальтонізму (колірної сліпоти). Російський лікар В.М.Флоринський у монографії ,,Вдосконалення і виродження людского роду” (1866 р.) писав , що збереження і покращення її можливе лише за доцільного підбору подружжя.
Генетика людини як наука виникла завдяки працям англійського вченого Ф.Гальтона (1822-1911 рр.). він разом із Г.Менделем є один із засновників генетики як науки. Гальтон вивчав успадкування розумових здібностей , обдарованість , таланту людини , створив особливий напрям генетики – євгеніку , призначення якої – вдосконалити людину і людський рід.
Менделісти поч. ХХ ст. (У.Бетсон , В.Іогенсен) та ін. Вивчали якісні ознаки, які визначаються окремими генами і стверджували , що ці гени визначають характер спадковості людини. У 1918 р. Фішер довів, одні ознаки людини визначаються якістю , тобто окремими генами, а другі – кількістю.
У 1900 р. К.Ланштейнер відкрив групи крові системи АВО і тим самим заклав початок вивченню поліморфних ознак людини.
У 1913 р. було описано поліморфізм відносно виявлення здібностей відчувати смак розчину фенілтіосечовини. Лікар А.Гаррод (1902 р.) розробляв проблему порушення обміну речовин у людини захворюванні алкаптанурією.
Алкаптанурія – спадкове захворювання , яке зумовлено неповноцінністю фермента
оксидази гемогентрозинової кислоти. А.Гаррод сформував знамените положення про спадкові дефекту обміну речовин , тобто заклав основи біохімічної генетики.
Дж.Бідл і Є.Теймен , вивчаючи біосинтез тіаміну , встановили , що за синтез
кожного ферменту відповідає певний ген. Вони виклали гіпотезу ,,один ген –один фермент” .
У 1908 р. Д.Харді , математик із кембріджського університету , і В.Вайнберг,лікар із Штутгарда , незалежно один від одного заклали основи популяційної генетики і сформулювали закон , який носить їхнє ім’я. Закон Харді-Вайберга було відкрито під час вивчення розподілу різних ознак у популяцій людини.
У 20-х і 30-х рр. такі досліди , як Р.Фішер і Дж.Халдейн в Англії , С.Вайт у США , Г.Дольберг у Швеції , Л.Хогбен і Ф.Берштейн у Німеччині внесли великий вклад теорію генетики і еволюції, в розробку статистичних методів вивчення генетики людини. Ці вчені описали методи аналізу закономірності успадкування, розщеплення , щеплення ознак і визначення частоти мутації.
У 1956 р. Д.Тійло і А.Леван встановили , що кількість хромосом у соматичних клітинах – 46 , після чого були виявлені зміни хромосом при різних захворюваннях.
У 1969 р. Т.Каперсон запропонував диференціальне фарбування хромосом , що
дало змогу розрізняти кожну з хромосом окремо і виявляти зміни їх.
В Україні питаннями медичної генетики займалися такі відомі вчені, як Т.І.Юдін Б.М.Манківський.
3. Особливості генетики людини. Дослідження генетики людини пов'язане з великими труднощами, причини - у неможливості експериментального схрещування, повільній зміні поколінь, малій кількості потомків у кожній сім'ї. Крім того, на відміну від класичних об'єктів, що вивчаються у загальній генетиці, у людини складний каріотип, велика кількість груп зчеплення. Проте, не зважаючи на всі ці труднощі, генетика людини успішно розвивається.
Неможливість експериментального схрещування компенсується тим, що дослідник, спостерігаючи широку людську популяцію, може вибирати із тисяч шлюбних пар ті, які необхідні для генетичного аналізу. Метод гібридизації соматичних клітин дозволяє експериментальне вивчати локалізацію генів у хромосомах, проводити аналіз груп зчеплення
При вивченні генетики людини використовуються такі методи: генеалогічний, близнюковий, популяційно-статистичний, дерматогліфичний, біохімічний, цитогенетичний, гібридизації соматичних клітин і метод моделювання.
У людини встановлені всі 24 теоретично можливі групи зчеплення генів: із них 22 локалізовані у аутосомах, у кожній із яких міститься по кілька сот генів. Більше 100 генів локалізовано у статевих хромосомах.
У ссавців, у тому числі і в людини, Х- і Y-хромосоми мають гомологічну ділянку, в якій відбувається їх синапсис і можливий кросинговер. Всі гени, які локалізовані у статевих хромосомах людини, можна поділити на три групи залежно від того, у яких ділянках статевих хромосом вони знаходяться.
Перша група охоплює гени, які локалізовані у тій частині X-хромосоми, що не має гомологічної ділянки у У-хромосомі. Вони повністю зчеплені зі статтю, передаються виключно через X-хромосому. До їх числа відносяться рецесивні гени гемофілії, дальтонізму, атрофії зорового нерва тощо. Домінантні гени із цієї ділянки однаково проявляються в осіб обох статей, рецесивні - у жінок тільки у гомозиготному, а у чоловіків - і гемізиготному стані.
Другу групу складає невелика кількість генів, які розташовані у непарній ділянці Y-хромосоми. Вони можуть зустрічатися тільки у осіб чоловічої статі і передаються від батька до сина. До них відносяться: волосатість вух, іхтіоз (шкіра у вигляді луски риби), перетинки між пальцями на ногах.
Третя група- гени, які розташовані у парному сегменті статевих хромосом, тобто гомологічному для Х- і Y-хромосом. їх називають неповно або частково зчепленими зі статтю. Вони можуть передаватися як з Х-, так і з Y-хромосомою і переходити з однієї до іншої у результаті кросинговера.
4. Методи вивчення спадковості у людини. Генеалогічний метод грунтується на простеженні якої-небудь ознаки у ряді поколінь з вказівкою родинних зв'язків між членами родоводу. Генеалогія, у широкому розумінні слова,- родовід людини.Генеалогічний метод був введений у науку в кінці XIX ст. Ф. Гальтоном. Суть його полягає у тому, щоб з'ясувати родинні зв'язки і прослідкувати наявність нормальної або патологічної ознаки серед близьких і далеких родичів у даній сім'ї.Генеалогічний метод є основною зв'язуючою ланкою між теоретичною генетикою людини і застосуванням її досягнень у медичній практиці.
Близнюковий метод - один з найбільш ранніх методів вивчення генетики людини, але він не втратив свого значення і сьогодні. Близнюковий метод дослідження був запропонований у 1876 р. англійським антропологом і психологм Ф. Гальтоном. Він виділив серед близнят дві групи: однояйцеві (монозиготні) і двояйцеві (дизиготні). На сьогодні в середньому на кожні 100 пологів приходиться одне народження близнят. Демографи розрахували, що на Землі проживає близько 50 млн пар близнят. Приблизно одну третину всіх близнят складають однояйцеві, а дві третини - двояйцеві. Кількість моно-зиготних близнят у різних регіонах земної кулі величина відносно постійна, з невеликими коливаннями. Наприклад. в Італії 0,37%, в Данії- 0,38, в Японії - 0,40, в США - 0,39, в Австралії - 0,38 %. Із наведених даних видно, що фактори, які впливають на появу однояйцевих близнят, майже не залежать від умов зовнішнього середовища.
У старших вікових групах народження дизиготних близнят зустрічається частіше. Дослідження показали, що певну роль у народженні близнят має спадкова схильність до багатоплідної вагітності. Відомі випадки повторного народження близнят у одній сім'ї. На протязі останніх десятиріч кількість близнят знижується, причому це зниження стосується переважно двояйцевих близнят. Монозиготні близнята розвиваються із роз'єднаних бластомерів, які утворилися після дробіння однієї заплідненої яйцеклітини і, отже, мають однакоііий генотип. Монозиготні близнята при нормальному ембріональному розвитку завжди однієї статі. У більшості випадків у них є одна плацента, проте не завжди можна зробити висновок про зиготність близнят на підставі наявності однієї чи двох плацент. Якщо розділення відбувається на протязі перших п'яти днів після запліднення, то кожний зародок буде мати власну плідну оболонку і плаценту. Цей варіант зустрічається приблизно у 25 % однояйцевих близнят. Якщо розщеплення відбувається на стадії розвиненої морули (5-12-й день), тоді однояйцеві близнята мають одну плаценту. Якщо ж процес розщеплення запізнюється і відбувається після ІЗ-15-го дня, то часто повного роз'єднання монозиготних близнят не відбувається і виникають різні деформації, зрощення і каліцтва. Прикладом може бути народження в Сіамі (теперішній Таїланд) у 1811 р. двох близнят хлопчиків, які зрослися грудьми. Згодом всі близнята, які зрослися якою-небудь частиною тіла, стали називатися сіамськими. Енг і Чанг прожили 63 роки, стан медицини в ті роки не давав можливості зробити операцію їх роз'єднання. Спроби роз'єднання зрослих близнят пов'язані з великим ризиком, проте на сьогодні відомі випадки вдалих операцій. Причини народження двояйцевих близнят - одночасне дозрівання двох і більше яйцеклітин. Це може статися у одному яєчнику або у обох. У деяких жінок бувають множинні овуляції. Певну роль в цьому відіграє спадкова схильність, але не можна відкидати і вплив навколишнього середовища. Народження трьох, чотирьох і більше дітей у людини трапляється рідко. У 1934 р. в Канаді народились 5 дівчаток, у 1974 р. в Гданську - 3 хлопчика і дві дівчинки, які нормально росли і розвивались. Відомий випадок народження 5 дітей в Японії у 1981 р. У 1980 р. в Італії у 28-річної жінки народилось 6 дітей (дві дівчинки і чотири хлопчика). З генетичної точки зору двояйцеві близнята подібні як звичайні сибси, але на відміну від останніх у них більша спільність факторів при внутрішньоутробному (пренатальному) і частково постнатальному періодах розвитку.
Близнюковий метод використовується у генетиці людини для того, щоб оцінити ступінь впливу спадковості і середовища на розвиток якої-небудь нормальної або патологічної ознаки. Оскільки у монозиготних близнят однакові генотипи, то наявні відмінності викликаються умовами середовища у період або внутрішньоутробного розвитку, або формування організму після народження. Великий інтерес для вирішення ряду питань мають випадки, коли партнери за якихось причин росли і виховувалися у різних умовах. Прояв конкордантності ряду фізіологічних ознак у такому випадку пояснюється впливом генотипу. З іншого боку, різнояйцеві близнята дозволяють проаналізувати інший варіант: умови середовища (коли близнята живуть поряд) однакові, а генотипи у них різні.
Метод дерматогліфіки. Дерматогліфіка (гр. derma-шкіра, qliphe-малювати) -це вивчення рельєфу шкіри на пальцях, долонях і підошвах. На відміну від інших частин тіла тут є епідермальні виступи - гребені, які утворюють складні візерунки. Ще у давнину у Китаї і Індії звернули увагу на те, що візерунки на пальцях і долонях строго індивідуальні, і користувалися відбитками пальців замість підписів. На землі немає людей з однаковими малюнками на пальцях (крім монозиготних близнят). У 1892 р. Ф. Гальтон запропонував класифікацію цих візерунків, яка дозволила використовувати метод для ідентифікації особи у криміналістиці. Таким чином, виділився один із розділів дерматогліфіки - дактилоскопія (вивчення візерунків на подушечках пальців). Інші розділи дерматогліфіки - пальмоскопія (малюнки на долонях) - плантоскопія (вивчення дерматогліфіки підошв).
Закладка візерунків відбувається між 10 і 19 тижнями внутрішньоутробного розвитку; у 20-тижневих плодів уже добре помітні форми візерунків розгалуження нервових волокон.Повне формування деталей будови дотикових візерунків завершується до шести місяців, після чого вони залишаються незмінними до кінця життя. При пошкодженні шкіри (опік, відморожування, травми) їх малюнок через деякий час повністю відновляється до деталей. Звичайно, відновлення можливе до тих пір, доки пошкодження не пов'язане з глибокою травмою, яка тягне утворення рубців із щільної сполучної тканини. Дерматогліфічні дослідження мають важливе значення у визначенні зиготності близнят, у діагностиці деяких спадкових хвороб, у судовій медицині, у криміналістиці для ідентифікації особи.
Біохімічні методи використовуються для діагностики хвороб обміну речовин, причиною яких є зміни активності окремих ферментів. За допомогою біохімічних методів відкрито близько 5000 молекулярних хвороб, які є наслідком прояву мутантних генів. При різних типах захворювання вдається або визначити сам аномальний білок - фермент, або проміжні продукти обміну. Ці методи дуже трудомісткі, вимагають спеціального обладнання і тому не можуть бути використані для масових популяційних досліджень з метою раннього виявлення хворих із спадковою патологією обміну.
Популяційно-статистичний метод дозволяє вивчати поширення окремих генів у популяціях людей. Звичайно проводиться безпосереднє вибіркове дослідження частини популяції або вивчають архіви лікарень, пологових будинків, а також проводять опитування шляхом анкетування. Вибір способу залежить від мети дослідження. Останній етап полягає у статистичному аналізі.
Одним з найбільш простих і універсальних методів є метод, запропонований Г. Харді і В. Вайнбергом (див. гл. II). Є і ряд інших спеціальних математичних методів. У результаті цього можна визначити частоту генів у різних групах населення, частоту гетерозиготних носіїв ряду спадкових аномалій і хвороб.
Досліджувані популяції можуть розрізнятися за біологічними ознаками, географічними умовами життя, економічним станом. Вивчення розповсюдженості генів на певних територіях показує, що їх можна розділити на такі категорії: 1) мають універсальну поширеність (до них відноситься більшість відомих генів); це рецесивні гени фенілкетонурії і деяких інших форм розумової відсталості, які зустрічаються у гетерозиготному стані у І % населення Європи; ген дальтонізму, який проявляється у 7 % чоловіків і 0,5 % жінок, але у гетерозиготному стані цей ген мають ІЗ % жінок; 2) зустрічаються локально, переважно у певних районах; наприклад, ген серпоподібно-клітинної анемії, який поширений у країнах Африки і Середземномор'я; ген, що зумовлює природжений вивих стегна, має високу концентрацію у корінного населення північно-східної частини Євразії.
Популяційно-статистичний метод дозволяє визначити генетичну структуру популяцій (співвідношення між частотою гомо- і гетерозигот). Нові можливості для проведення генетичного аналізу відкриває використання електронно-обчислювальної техніки. Знання генетичного складу популяцій населення має велике значення для соціальної гігієни і профілактичної медицини.
Патогенетичний метод. Принципи цитогенетичних досліджень сформувалися на протязі 20-30-х років на класичному об'єкті генетики - дрозофілі і деяких рослинах. Метод грунтується на мікроскопічному дослідженні хромосом.
Нормальний каріотип людини включає 46 хромосом, із них 22 пари ауто-сом і 2 статеві хромосоми. До 1956 р. кількість хромосом у людини не була точно встановлена, це вдалося шведським вченим Д. Тийо і А. Левану. Для ідентифікації хромосом застосовують кількісний морфометричний аналіз. З цією метою проводять вимірювання довжини хромосоми у мікрометрах. Визначають також співвідношення довжини короткого плеча до довжини всієї хромосоми (центромерний індекс).
5. Мутації та їхні прояви у фенотипі людини. Поняття про спадкові хвороби. У людини, як і у інших хромосомні хвороби, а захворювання, які зв'язані з мутаціями на молекулярному рівні, називають генними хворобами.
Завданням медичної генетики є виявлення і профілактика спадкових хвороб. В останні десятиліття значно змінилась структура захворювань населення.Захворювання з екзогенними факторами етіології , як то інфекційні авітамінози, отруєння , відійшли на задній план завдяки значним досягненням мікробіології , імунології і біохімії , а на перший план виступили захворювання з ендогенними факторами етіології , тобто спадкові.
За даними експертів Всесвітньої організації охорони здоров’я (ВООЗ) , одна дитина із 100 новонароджених страждає важким спадковим захворюванням внаслідок уродження хромосом , у 4% дітей спостерігаються різні генетичні дефекти. Генетичні дефекти є також причиною 40% спонтанних абортів. Кожна людина є носієм 15-20 потенційно дефектних генів. Розроблення сучасних біохімічних , цитологічних і генетичних методів досліджень сприяло розкриттю молекулярної сутності багатьох захворювань. Було встановлено , що в розвитку як спадкових , так і не спадкових (екзогенних)захворювань істотне значення має стан генетичного апарату клітин організму.
Сьогодні генетика є базовою для всіх біологічних наук , у тому числі й медичних.
Завданням сучасної медицини є поступовий перехід із сфери лікування хворих у
сферу запобігання хворобам і збереження здоров’я населення.
Г
 енетичні
хвороби— порушення
нормальної роботи організму
через порушення послідовності генів
або структури хромосом.
Тоді як деякі хвороби, наприклад рак,
виникають через генетичні порушення,
придбані кількома клітинами протягом
життя, термін «генетична хвороба»
зазвичай посилається на наявність
хвороб у всіх клітинах тіла і присутніх
з моменту зачаття. Деякі генетичні
хвороби викликаються хромосомними
порушеннями, що виникають через помилки
процесу мейозу
між генеративними клітинами— спермою
і якцеклітиною.
Приклади таких генетичних хвороб
включають
енетичні
хвороби— порушення
нормальної роботи організму
через порушення послідовності генів
або структури хромосом.
Тоді як деякі хвороби, наприклад рак,
виникають через генетичні порушення,
придбані кількома клітинами протягом
життя, термін «генетична хвороба»
зазвичай посилається на наявність
хвороб у всіх клітинах тіла і присутніх
з моменту зачаття. Деякі генетичні
хвороби викликаються хромосомними
порушеннями, що виникають через помилки
процесу мейозу
між генеративними клітинами— спермою
і якцеклітиною.
Приклади таких генетичних хвороб
включають
синдром Дауна (додаткова хромосома 21)
 Розумова
та фізична відсталість,
Розумова
та фізична відсталість,
Напіввідкритий рот
Монголоїдний тип обличчя. Косо
розміщені очі.
Широке перенісся
Стопи і кисті короткі та широкі, пальці неначе обрубані
 Пороки
серця
Пороки
серця
Тривалість життя знижується у 5-10 разів
синдром Тернера (45X0)(2n = 45 хромосом - відсутня одна статева хромосома (Х0).
Спостерігається у дівчаток
Порушення пропорцій тіла (низький ріст, укорочені ноги, широкі плечі, коротка шия, схильність до ожиріння)
Розумова відсталість
Криловидна шкірна складка на шиї
Пороки внутрішніх органів
Недорозвинутість статевих органів, безпліддя.)
синдром Кляйнфелтера (чоловік з 2 X-хромосомами).
2n = 47,48,49 хромосом – 44 + ХХУ; 44 + ХХХУ; 44 +ХУУ; 44 + ХХУУ
Спостерігається у хлопців
Високий ріст, порушення пропорцій тіла (довгі кінцівки, вузька грудна клітка)
Відсталість у розвитку
Недорозвинутість статевої системи, безпліддя
Жорстокі
Інші генетичні зміни можуть відбуватися протягом формування генеративних клітин одним з батьків. Один приклад — мутації тріплетного повторення, які можуть викликати хворобу Хантінгтона. Дефектні гени також можуть бути цілком успадковані від батьків. В даному випадку, генетична хвороба називається спадковою хворобою. Така хвороба часто виникає несподівано, коли два здорові носії дефектного рецесивного гена передають дитині дві дефектні копії гена, але також трапляються, коли дефектний ген є домінантним.
У каріотипі нормальної жінки є дві X-хромосоми, і одна із них утворює тільце статевого хроматину. Статевий хроматин можна визначити у будь-яких тканинах. Частіше всього досліджуються епітеліальні клітини слизової оболонки щоки (буккальний зскрібок). Це особливо зручно при масових дослідженнях. Кількість тілець статевого хроматину у людини та інших ссавців на одиницю менша, ніж число Х-хромосом у даної особини. У жінок, які мають каріотип ХО (моносомія-, синдром Шерешевського - Тернера), ядра клітин не мають статевого хроматину. При синдромі трисомії -X у жінки утворюються два тільця, у чоловіка з каріотипом 47 (ХХУ) - є одне тільце (як у нормальних жінок).
Статевий хроматин можна визначити і на мазках крові, у ядрах нейтрофілоцитів; вони мають характерний вигляд барабанних паличок, які відходять від складно-дольчастого ядра цих лейкоцитів. У нормі у жінок ці структури виявляються у 3-7 % нейтрофілоцитів, у чоловіків вони взагалі відсутні. Деякі автори вважають, що цей метод більш достовірний, ніж буккальний зскрібок, але внаслідок великої трудомісткості він використовується тільки при спеціальних дослідженнях.
Визначення статевого хроматину використовують і у судовій медицині, коли необхідно за плямами крові встановити статеву належність, при аналізі, коли необхідно встановити, чоловікові чи жінці належить знайдена частина трупа, навіть через тривалий термін після смерті.
Виявлення Y-хроматину впроваджується у практику медико-генетичних консультацій.
Успадкування резус-фактора. У макак-резус із еритроцитів у 1940 р. виділено антиген, який назвали резус-фактором (Rh-фактор). Згодом він був знайдений і у людей. Близько 85 % європейців його мають, тобто є резус-позитивними (Rh+), а у 15% резус-негативних (Rh-) він відсутній.
У нормі в осіб з резус-негативною кров'ю не виробляються антитіла до резус-фактора, але вони почнуть вироблятися у результаті переливання резус-позитивної крові як захисна реакція проти чужорідного антигена.
Успадкування резус-фактора зумовлене трьома парами генів -С, D, К, які тісно зчеплені між собою, тому практично успадкування його частіше всього імітує моногенне успадкування.
Резус-позитивний фактор зумовлений домінантними генами. При шлюбі жінки з резус-негативною кров'ю і чоловіка з наявністю резус-фактора за умови гомозиготності батька всі діти будуть резус-позитивними, а при гетерозиготності буде спостерігатися розщеплення у відношенні 1 : 1.
Якщо у жінки з резус-негативною кров'ю дитина, що народиться, успадкує резус-фактор, перша вагітність може завершитись цілком нормально. Але при цьому у кров'яному руслі матері утворюються антитіла до Rh+-фактора. При наступній вагітності ці антитіла проникають у кров плода і викликають руйнування еритроцитів, які мають антиген Rh+. З кожною наступною вагітністю, несумісною за антигенами, кількість антитіл до Rh+-фактора у тілі матері зростає. Іноді гинуть недоношені ембріони, спостерігається мертвонародження. У зв'язку з прониканням у кров'яне русло дитини антитіл у неї розвивається гемолітична хвороба, що призводить до руйнування еритроцитів. Врятувати новонародженого може тільки термінове переливання крові з повною її заміною.
Із сказаного також має бути зрозумілим, що для переливання крові необхідно досліджувати її на Rh-фактор. Переливання несумісної за цим фактором крові дівчатам і жінкам зовсім недопустиме, бо може викликати безпліддя.
Самостійне вивчення 4
Етологічна структура популяції - це система взаємозв'язків між: особинами, яка проявляється в поведінці. Науку про біологічні основи поведінки тварин називають етологією (від грец. етос - характер, норов і логос). Особинам різних видів властивий поодинокий або груповий спосіб життя. У першому випадку особини популяції більш-менш розділені просторово і збираються разом лише на період розмноження, міграцій тощо (скорпіони, тетеруки, качки-крижні та ін.). Груповий спосіб життя пов'язаний з утворенням постійних родин, колоній, зграй, табунів тощо. Спільне існування організмів у вигляді постійних груп сприяє кращо-му пристосуванню до умов довкілля (захист від хижаків, ефективне полювання, виживання молоді тощо). Зазвичай у цих угрупованнях кожна особина займає певне положення (ранг), яке визначає поведінку, черговість доступу до їжі, участі (або неучасті) у розмноженні тощо.
Одиночний і сімейний спосіб життя.
Повністю одиночне існування організмів у природі не зустрічається, оскільки не була б можливою така основна функція будь-якого організму, як розмноження.
Приклад 1. У бурого ведмедя пари утворюються лише на короткий строк. Ведмеді-самці — «вітрогони». Навесні в популяції ведмедів починається гон, тобто шлюбний період. Самці знаходять і переслідують самок. Між «кавалерами» нерідко відбуваються жорстокі бої. «Весілля» триває 1—1,5 місяця, а після здійснення таїнства партнер і партнерша розходяться. Самець ніякої участі у ство-ренні сім'ї і вихованні свого потомства не бере. Через 7 місяців в барлозі з'являються ведмежата.
Приклад 2. Восени самці крижнів одягають «шлюбне» яскраве вбрання. На місцях зимівель птахи утворюють пари. Самець ретельно доглядає свою «наречену». Ранньої весни крижні повертаються додому і відразу приступають до будівництва гнізда. Мостять гнізда на непрохідних болотах на землі або на купинах. Після того як самка знесе 7—9 яєць і сяде на гніздо, самець її назавжди покидає.
Приклад 3. Поля ще вкриті снігом, тільки-но з'явилися перші таловини, а вже чути дзвінку, життєрадісну пісню польового жайворонка. Чудовим співом самець зачаровує самку, утворюється пара. Удвох мостять гніздо на землі. Самка насиджує яйця 14 днів. А самець у цей момент підгодовує її і співає свої чудові пісні. Коли з'являються пташенята, пісні припиняються і обидва батьки вигодовують молодь. Після вильоту молодих птахів із гнізда пари розпадаються.
Приклад 4. Морж — тварина полігамна. У «гаремі» самця може бути до 5 і більше самок, яких він ретельно охороняє. Така група утворюється лише на один сезон, а на наступний все починається спочатку.
Висновок 1. У видів з одиночним способом життя утворюються тимчасові пари, спільне існування яких зумовлене еволюційно закріпленими статевими інстинктами та умовами існування.
Для того щоб пара утворилася, природа надала самцям цілий ряд пристосувань: від характерного забарвлення і розмірів до специфічної ритуальної поведінки.
Приклад 5. Вибір партнерів у тварин супроводжується особливою шлюбною поведінкою. У павуків стрибунців самці домагаються парування із самкою після своєрідного танцю, протягом якого вони описують біля неї півкола, дедалі звужуючи їх, і набувають особливих поз, а наблизившись, починають швидко крутитися, захоплюючи самку. Така поведінка самців гальмує інстинкт агресії в самок, які реагують на них як на здобич. Загалом поведінка самок і самців у павуків буває різною: у багатьох всеїдних видів самець після спарювання відразу ж стає жертвою самки, у бігаючи — поспіхом тікає, у стрибунців — самка і самець розходяться спокійно, а в деяких видів якийсь час мирно уживаються в одному гнізді.
Приклад 6. Ми всі звикли, що боротьба за самку між самцями — це типовий приклад існування того чи іншого виду. Але іноді трапляються випадки, коли самки оспорюють право на самця. У популяції вимираючого виду тигра амурського саме між самками відбуваються жорстокі бої за право володіння самцем.
Деякі види існують у природі сім'ями. Сімейні пари лебедів, журавлів, голубів, вовків зберігаються на довгі роки. Такий спосіб життя посилює інстинкти, пов'язані з турботою про потомство.
Приклад 7. Вовки живуть сім'ями, пари в них постійні, часто на все життя. Лише смерть партнера змушує вовків шукати собі іншої napи. Сім'я звичайно невелика й складається, з батьків та вовченят поточного року, до яких іноді приєднуються минулорічні вовченята або вовки-одинаки. У захисті лігва, догляді за вовченятами, добуванні їжі беруть участь обидва батьки.
Приклад 8. У жаб турбота про потомство має велике значення. Вона спостерігається в тих видів, які не мають відповідного місця для відкладання яєць. Своєрідний спосіб турботи про потомство спостерігається в ринодерми Дарвіна. Тут про потомство дбає самець. Він запліднює відкладені самкою яйця, вміщує їх у рот і проштовхує в горловий мішок, де вони і розвиваються.
Приклад 9. Імператорські пінгвіни — моногами. У них зразкова сім'я. У розпалі антарктичної зими самка відкладає яйце. Самець тримає його на лапах, прикривши зверху складкою шкіри. Адже покласти його нікуди — навколо сніг. Самка в цей час іде зі своїми подругами на берег океану запасати їжу для майбутнього потомства, нагулюють жир. Через 2 місяці самки урочистою
процесією повертаються до залишених самців, які весь цей час голодували, а на світ уже з'явилися пінгвінята.
Приклад 10. Леви живуть особливими сім'ями — прайдами. Прайд складаєть-ся з самця-голови, самок (їх кілька) і молодих левів (останніх трьох років поносу). Коли молодому самцю виповнюється три роки, він покидає прайд або вступає в боротьбу за керівництво в ньому з батьком. Іноді в цьому йому допомагають інші молоді самці. Нерідко це може закінчитися трагічно. Полюванням займаються левиці, які проводять в одному прайді все своє життя. Вони годують левенят (будь-яка самка готова годувати будь-яке левеня) і доглядають їх.
Розрізняють сім'ї батьківського, материнського й мішаного типу залежно від того, хто з батьків бере на себе догляд за потомством.
Висновок 2. При сімейному способі життя територіальна поведінка тварин виявляється найяскравіше: різноманітні сигнали, маркірування, ритуальні форми погрози і пряма агресія забезпечують володіння ділянкою, достатньою для вигодовування потомства.
Більш ускладнені поведінкові зв'язки характерні для популяцій тварин, які живуть великими групами.
Груповий спосіб життя.
Тип групового способу |
Приклад |
Колонії— це групові поселення осілих тварин. Вони можуть існувати довго або виникати лише на період розмноження |
Найскладніший тип колонії у громадських комах — термітів, бджіл, мурашок |
Зграї — це тимчасові об'єднання тварин, які виявляють біологічно корисну організованість дій |
Зграї поділяють на дві категорії: 1) еквіпотенціальні, без вираженого домінування окремих членів (морські риби, сарана) 2) зграї з лідерами (вовки, гієни, койоти) |
Стадо — тривале об'єднання кочових тварин. Основу групової поведінки тварин у стадах становлять взаємовідносини домінування — підлеглості, що ґрунтуються на індивідуальних відмінностях між особинами |
Розрізняють групи з тимчасовим лідером (північні олені) та з вожаками. Вожаки керують стадом, у якому виникає розподіл прав — ранги (павіани, зебри). Ієрархія відносин у такому стаді досить складна |
Група — це невелике об'єднання кількох сімей |
Групами живуть річкові бобри, ондатри, горбаті кити, шимпанзе |
Поняття про ефект групи.
Життя в групі через нервову і гуморальну системи позначається на стані багатьох фізіологічних процесів в організмі тварин. Оптимізація фізіологічних процесів, яка веде до підвищення життєздатності при спільному існуванні, отримала назву ефект групи. Він виявляється як психофізіологічна реакція окремої особини на присутність інших особин свого виду. В овець поза стадом частішає пульс і дихання, а як тільки тварина побачить отару, ці процеси відразу ж нормалізуються. Кажани, які зимують поодинці, відзначаються вищим рівнем обміну речовин, ніж у колонії. Це призводить до підвищеної витрати енергії, виснаження і часто закінчується загибеллю тварини.
Ефект групи виявляється в прискоренні темпів росту тварин, підвищенні плодючості, швидшому виникненні умовних рефлексів, збільшенні середньої тривалості життя особини тощо.
Разом з тим, позитивний ефект групи виявляється лише до деякого оптимального рівня щільності копуляції. Якщо тварин стає надто багато, то це загрожує для всіх нестачею ресурсів середовища.
Особливості поведінки людини.
Людина розумна (Homo sapiens) – це результат біологічної еволюції, її ускладнення і видо-змінення. Однак відмінності людини від тварини досить істотні. Тварина, народжуючись на світ, має певний набір інстинктів, які обумовлюють стереотипи поведінки. Тобто вона в межах біологічних закономірностей генетично запрограмована на певну поведінку. Структура організму тварини, що виробилася в процесі біологічного розвитку, детермінує і потреби, і поведінку, забезпечуючи виживання за певних умов. У людини ситуація зовсім інша. Людство розвивається на протязі 40 тисяч років, відноситься до одного соціального виду (Homo sapiens), але тим не менше виявляє значну різноманітність в процесі поведінки. К.Маркс прокоментував цю ситуацію так: “Людина може вести себе за мірками будь-якого виду”. Серед представників Homo sapiens присутня індивідуальна варіативність поведінки, яка невідома тваринному світу. Біологічні передумови поведінки тварин зафіксовані в молекулах ДНК (генетичному коді). В умовах культурної еволюції інформація передається наступним поколінням шляхом наслідування, навчання і оволодіння знаннями і навичками. Поведінку людей визначають членоподільна мова, показ і приклад. Функції природжених інстинктів, які притаманні тваринам, у людини замінюється нормами (правилами) і спадкоємністю в поведінці. Людина радикальна відрізняється від тварини способом відношення до зовнішньої природи. Це відбито вже в біологічній специфіці людської істоти, де з’єднана як тваринна, так і людська природа. Вона є матеріальним синтезом “органічного тіла” людини і історії людської праці. Тому ступінь людської своєрідності по відношенню до тварини проявляється саме в людській діяльності. Якщо тварина не відрізняє себе від своєї життєдіяльності, то людина робить саме свою життєдіяльність предметом своєї волі і своєї свідомості. Виділення людини з природи відбулося завдяки: 1) вмінню виробляти знаряддя праці;
2) володінню членоподільною мовою;
3) наявності свідомості і розуму;
4) здібності бути моральною, підніматися над своїми природними
потягами і здійснювати вільний вибір. З іншого боку, людина істота соціальна, бо вона перетворює природні об’єкти, тобто займається виробництвом, предметно-практичною діяльністю. Саме в ході цієї діяльності людина формується як соціальна істота. Процес переходу від суто біологічних закономірностей розвитку людини як виду до культурних закономірностей її існування окреслюється досить широкими хронологічними рамками, у межах яких антропогенез, що розпочався близько 3 млн. років тому, доповнений соціогенезом (близько 1,6 млн. років тому) завершився появою Homo sapiens ( 35-40 тис.років тому). Інтенсивний розвиток пралюдини розпочався у льодовиковому періоді, що характеризувався різкою зміною кліматичних умов та природними катаклізмами. Це зрозуміло, змушувало гомінідів шукати засоби пристосування до нових умов і, найбільшою мірою, - цілеспрямовано змінювати обставини відповідно до потреб життя. Вихідними умовами такого типу пристосування були: розвинута вища нервова система, достатня для подальшої інтенсифікації інтелекту, здатність спілкуватися й використовувати підручні знаряддя у процесі пристосування, спільний спосіб існування. Тривалий груповий відбір цього виду гомінідів у подальшому сприяв розвиткові та вдосконаленню цих генетичних властивостей як засобів уже діяльного, а не суто органічного пристосування до навколишнього середовища. Діяльне пристосування тісно пов’язане з виникненням виробничої діяльності для створення засобів такого пристосування, що кваліфікується науковою як початок виходу пралюдей із тваринного стану. Виробниче-знаряддєва діяльність – вже людська характеристика, що відповідає культурно-еволюційним закономірностям. Вона є початком формування людських рис, бо потребує мислення, знань, мови як засобу передання життєво корисної інформації й відповідно – специфічної людської морфологічної інформації.
За свого виникнення виробнича діяльність була ще досить тваринною, близькою до умовно-рефлекторної поведінки. До неї вдавалася ще зоологічно організована спільнота. Основою вивільнення виробничої діяльності з тваринної оболонки став кооперативний механізм ( суто культурно-еволюційна закономірність) як оптимальна умова приборкання біологічних інстинктів ( зокрема, зоологічного індивідуалізму) та використання колективних зусиль для індивідуального виживання. Цей народжуваний культурною еволюцією механізм взаємодії став згодом засобом перетворення первісного стада в людський соціум ( родову общину), а далі – в суспільство як культурну спільноту.
Нові способи колективного існування зумовлювали швидке зростання інтелектуальних здібностей пралюдей, ускладнення їхніх навичок, розширення кола їхніх знань. Виживання виду починало більше залежати не від силової переваги у внутрішньовидовій боротьбі, а від володіння життєво необхідною інформацією. Більше того виробничо-діяльне існування пралюдей все очевидніше суперечило механізмові біологічних відносин. Адже зміст знань і зростання їхнього обсягу потребували нових методів оволодіння, нагромадження й передання їх. Нагромаджена інтенсивною роботою мозку життєво необхідна інформація вже не може бути генетично засвоєною та переданою, бо якість, кількість і швидкість отримання інформації тепер не збігається з умовами генетичного кодування. Виникає потреба в пам’яті, яка не пов’язана з генетичним механізмом. Вже на рівні вищих ссавців поряд із генетичним існує наслідування за принципом “роби як я!” Цей первинний механізм негенетичного наслідування дозволяє передавати наступним поколінням сприятливі навички поведінки, він розрахований на найпростішу інформацію, яку можна засвоїти шляхом імітації. Більш складна і розширене інформація потребувала винайдення засобу специфічно культурного кодування – мови, системи цілеспрямованого навчання. Тому культурно-еволюційний розвиток сприяє формуванню більш довершеної, більш універсальної форми пам’яті. Вона могла реалізувати лише в умовах гарантованої безпеки для носіїв знань. Постала необхідність докорінної перебудови всієї системи відносин у первісних спільнотах, яка вимагала відмови від суто біологічного способу, вдосконалення виду –внутрішньовидової боротьби. Протягом багатьох сотень років відбувалося поступове обмеження біологічних інстинктів через формування системи заборонних форм поведінки для кожного члена спільноти – як вираз волі праобщини стосовно індивідуальних обов’язків у процесі спільного виживання. Народжувалась ще одна засада культурної еволюції – мораль. Завдяки цьому відбір на біологічному рівні було витіснено відбором на рівні організації соціальних спільнот. Розвиток людини як біологічної істоти закінчується у кінці мезоліту – на початку неоліту. З точки зору морфологічної будови вона відповідає сучасним стандартам: за способом існування повністю вивільнилася з біологічно-інстинктивної форми, всі біологічні інстинкти взяті під соціальний контроль. Людина остаточно перейшла у стадію культурно-еволюційного розвитку. Визначення принципів періодизації 40-тисячолітньої культурної еволюції є дуже складною і дискусійною проблемою. Існує чимало варіантів цієї періодизації. Вони вже давно й активно “працюють” у науці, що не заважає появі і апробації нових підходів до проблеми.
Самостійне вивчення 5
Поява людини як біологічного виду якісно відрізняється від процесів виникнення нових видів у тварин. Найважливіша якісна відміна полягає в тому, що еволюція людини відбувалася під впливом не тільки біологічних, а й соціальних чинників. На ранніх етапах антропогенезу в найдавніших і давніх людей провідну роль для їхньої еволюції відіграв природний добір.
Природний добір- це складний історичний процес у живій природі, що полягає в збереженні організмів з корисними в даних умовах індивідуальними змінами і зникненні непристосованих.
Проте вже навіть на цих етапах добір відбувався не тільки за фізичними показниками (сила, витривалість тощо) людини, а й за рівнем її інтелектуального (розумового) розвитку (уміння виготовляти найпростіше знаряддя праці, використання їх, здатність до спілкування з іншими членами племені, навички колективних дій тощо). Отже, природний добір сприяв збереженню особин з вираженими соціальними рисами. З удосконаленням трудової діяльності й мови наших предків ця спрямованість добору посилювалася. Особини, які були здатні винаходити нові знаряддя, навчатися й аналізувати обставини, мали більше можливостей вижити. У процесі еволюції люди почали створювати штучне середовище для життя: опалювані житла, одяг, запаси продуктів. Тому залежність людей від несприятливих чинників поступово зменшувалася, а отже, зменшувалося значення в їхньому житті природного добору.
Головними соціальними чинниками, що вплинули на еволюцію людини, стали праця, життя в суспільстві, членороздільна мова та абстрактне мислення. Праця- це планова діяльність, пов'язана з використанням спеціальних знарядь.
Зміни, що відбулися в будові руки первісної людини, сприяли появі можливості виготовляти прості знаряддя праці. Цей процес тривав сотні тисяч років. З часом знаряддя праці вдосконалювалися. Водночас і рука людини, яка їх створювала, сама стала знаряддям праці.
Важливе значення для становлення людини мав суспільний характер праці. Спільне полювання, підтримування вогню, виготовлення знарядь праці зумовили необхідність спілкування членів племені і виникнення потреби передавати інформацію. Так поступово розвинулася членороздільна мова як спосіб передачі інформації від однієї людини до іншої, а також знаряддя для передавання досвіду від покоління до покоління.
Розвиток мови вплинув на подальший розвиток головного мозку та процеси мислення. У предків сучасної людини розвинулася здатність до абстрактного мислення, тобто до виокремлення істотних ознак у явищах і тілах природи, їх порівняння, встановлення логічних зв'язків, узагальнення. Це стало підґрунтям для розвитку інтелекту людини, який досяг неймовірного втілення в різноманітних винаходах.
Вид Людина розумна має чітко виражені зовнішні ознаки, що історично виникли в період її активного розселення по планеті Земля та є наслідком пристосування до конкретних умов навколишнього середовища. Оскільки ці умови (клімат, особливості місцевості, температурний режим тощо) істотно відрізнялися, то в цілих групах людей, пов'язаних з певними районами, виникли специфічні зовнішні ознаки (відповідний колір шкіри, тип і колір волосся, форма носа, губ, колір і розріз очей тощо). Це сприяло виникненню рас.
Раса- це група людей, що має спільне походження і характеризується комплексом відповідних спадкових ознак, які сформувалися як пристосування до різних умов середовища впродовж багатьох поколінь.
У наш час учені виділяють три великі раси виду Людина розумна - європеоїдну (євразійську), австрало-негроїдну, монголоїдну (азіатсько-американську). Кожна з них має ще низку підрас.
Європеоїдна раса поширена в Європі, Передній Азії, Північній Африці й Північній Індії. Люди цієї раси мають світлу або смугляву шкіру, пристосовану до проникнення ультрафіолетових променів, пряме або хвилясте найчастіше світле волосся, вузький продовгуватий ніс, що сприяє зігріванню холодного повітря, тонкі губи, великі голубі або сірі очі.
Монголоїдна раса поширена в Азії, Північній і Південній Америці. Для людей цієї раси характерні світла шкіра, пристосована до північного клімату, оскільки може забезпечити в організмі синтез вітаміну Б в умовах малої інтенсивності сонячного світла, пряме темне волосся, широкий ніс, тонкі губи, вузький розріз очей і шкірна складка у внутрішньому куті ока (пристосування до постійних вітрів степу, що несуть пісок і пил).
До австрало-негроїдної раси, поширеної в Африці, Центральній і Південній Америці, належать також аборигени Австралії. Характерним для цієї расової групи є специфічний чорний колір шкіри, що захищає тіло від ультрафіолетових променів в умовах інтенсивного сонячного світла, кучеряве густе чорне волосся, широкий ніс, товсті губи, великі темні очі.
За основними ознаками, характерними для виду Людина розумна (об'ємом мозку, будовою кисті та стопи, формою хребта, будовою голосових зв'язок, інтелектуальними здібностями), раси абсолютно рівноцінні і перебувають на одному й тому самому рівні еволюційного розвитку. Це спростовує вигадки ідеологів расизму про «вищі» раси та нації. Усе сучасне людство - це один біологічний вид - Homo sapiens.
ЛЮДИНА
РОЗУМНА