

Билет №33.
1 Вопрос. Витамин B6 -- общее название трёх веществ: пиридоксина, пиридоксаля, пиридоксамина, а также их фосфатов, среди которых наиболее важен пиридоксальфосфат.В пище человека встречаются пиридоксин, пиридоксаль и пиридоксамин, а также их фосфаты. В человеческом организме любое из этих веществ превращается в пиридоксальфосфат.
Витамин B6 (пиридоксин, пиридоксаль, пиридоксамин) содержится во многих продуктах. Особенно много его содержится в зерновых ростках, в грецких орехах и фундуке, в шпинате, картофеле, моркови, цветной и белокочанной капусте.. Витамин B6 (пиридоксин, пиридоксаль, пиридоксамин) синтезируется в организме кишечной микрофлорой. Пиридоксин представляет собой бесцветные кристаллы, растворимые в воде. В большинстве съедобных растений пиридоксина нет, или он встречается в незначительных количествах. Но существуют растения, например, питайя, исключительно богатые пиридоксином. Пиридоксин синтезируется некоторыми бактериями. Также он содержится в мясных и молочных продуктах, но он менее устойчив к высоким температурам, чем другие формы витамина B, поэтому в варёных и жареных мясных продуктах его мало. Вегетарианцы могут получить пиридоксин из кожицы некоторых овощей.
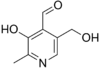
Пиридоксаль.
Брутто-формула пиридоксаля C8H9NO3.
Пиридоксаль отличается от пиридоксина
тем, что вместо одной из трёх гидроксогрупп
к пиридиновому кольцу присоединена
карбонильная группа, так что пиридоксаль
является одновременно и альдегидом.
Пиридоксамин
отличается от пиридоксина тем, что
вместо одной из трёх гидроксогрупп к
пиридиновому кольцу присоединена
аминогруппа. Пиридоксамин содержится
в мясе животных.

В
тканях все три формы витамина активно
превращаются в кофермент - пиридоксальфосфат
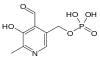 (ПФ), необходимый для продукции энергии
из аминокислот и поэтому рассматривается
как энергореализующий витамин. Причинами
развития недостаточности пиридоксина
могут быть хронические заболевания
желудочно-кишечного тракта, а также
наследственные дефекты в функционировании
пиридоксинзависимых ферментов. Состояние
недостаточности пиридоксина возникает
у женщин, принимающих противозачаточные
средства. Причиной этого являются
эстрогены, а не прогестерон. Более низкий
по сравнению с нормой уровень пиридоксина
у этих женщин вызывает сонливость,
слабость, умственную заторможенность
и ухудшение обмена веществ.
(ПФ), необходимый для продукции энергии
из аминокислот и поэтому рассматривается
как энергореализующий витамин. Причинами
развития недостаточности пиридоксина
могут быть хронические заболевания
желудочно-кишечного тракта, а также
наследственные дефекты в функционировании
пиридоксинзависимых ферментов. Состояние
недостаточности пиридоксина возникает
у женщин, принимающих противозачаточные
средства. Причиной этого являются
эстрогены, а не прогестерон. Более низкий
по сравнению с нормой уровень пиридоксина
у этих женщин вызывает сонливость,
слабость, умственную заторможенность
и ухудшение обмена веществ.
2 Вопрос. Цикл трикарбоновых кислот — центральная часть общего пути катаболизма, циклический биохимический аэробный процесс, в ходе которого происходит превращение двух- и трёхуглеродных соединений, образующихся как промежуточные продукты в живых организмах при распаде углеводов, жиров и белков, до CO2. При этом освобождённый водород направляется в цепь тканевого дыхания, где в дальнейшем окисляется до воды, принимая непосредственное участие в синтезе универсального источника энергии — АТФ.
Цикл Кребса
— это ключевой этап дыхания всех клеток,
использующих кислород, центр пересечения
множества метаболических путей в
организме. Кроме значительной
энергетической роли циклу отводится
также и существенная пластическая
функция.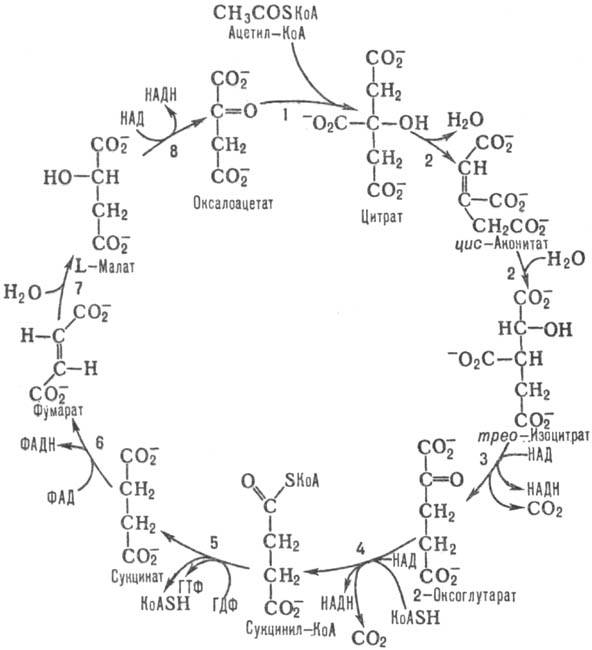 В реакции 1 цикла, катализируемой цитрат
- оксалоацетатлиазой, CH3C(O)SKoA стереоспецифично
конденсируется с карбонильной группой
оксалоацетата с образованием цитрата
и свободного KoASH. Реакция сопровождается
значительным изменением свободной
энергии и является практически
необратимой. Активность митохондриального
фермента у дрожжей ингибируется АТФ.
В реакции 1 цикла, катализируемой цитрат
- оксалоацетатлиазой, CH3C(O)SKoA стереоспецифично
конденсируется с карбонильной группой
оксалоацетата с образованием цитрата
и свободного KoASH. Реакция сопровождается
значительным изменением свободной
энергии и является практически
необратимой. Активность митохондриального
фермента у дрожжей ингибируется АТФ.
Реакция 2 цикла, катализируемая аконитатгидратазой, изомеризация цитрата в изоцитрат путем последовательной дегидратации - регидратации через промежут. образование цис-аконитата. Реакция обратима, равновесие сдвинуто в сторону синтеза цитрата, однако в условиях непрерывного функционирования цикла конечным продуктом реакции является изоцитрат.
В реакции 3, катализируемой НАД- или НАДФ-зависимой изоцитратдегидрогеназой, происходит дегидрирование изо-цитрата при атоме С-2 с одновременным декарбоксилированием и образованием 2-оксоглутарата и СО2. Бактерии содержат НАДФ-зависимую изоцитратдегидрогеназу, активность которой регулируется хим. модификацией - фосфорилированием (инактивация) и дефосфорилированием (активация) при участии бифункционального фермента изоцитратдегидрогеназа-киназа (фосфатаза). Эукариоты содержат обе формы изоцитратдегидрогеназы. Активность НАДФ-зависимого фермента, локализованного в митохондриальном матриксе и цитозоле, контролируется продуктами реакции.
Реакция 4 катализируется мультиферментным 2-оксоглутаратдегидрогеназным комплексом, состоящим из трех осн. ферментов: 2-оксоглутаратдегидрогеназы, дигидролипоилсукцинилтрансферазы и дигидролипоилдегидрогеназы.
Сукцинил-КоА в реакции 5, катализируемой сукцинил-КоА-синтетазой, подвергается распаду, в результате которого энергия тиоэфирной связи сукцинил-КоА запасается в виде синтезированного нуклеозидтрифосфата.
В реакции 6, катализируемой сукцинатдегидрогеназой, происходит превращение сукцината в фумарат. Фермент входит в состав более сложного сукцинатдегидрогеназного комплекса
Фумаратгидратаза, катализирующая реакцию 7, осуществляет гидратирование фумарата с образованием L-малата.
Последняя реакция цикла, 8, катализируется L-малатдегидрогеназой.
3 Вопрос. Желтуха – это заболевание, характеризующееся желтой окраской кожи и слизистых в результате накопления билирубина. Основная причина этого явления – гипербилирубинемия. Причинами гипербилирубинемии могут быть:
· усиление гемолиза эритроцитов и увеличение образования билирубина, превышающее способность печени экскретировать его;
· повреждение печени, приводящее к нарушению секреции билирубина в желчь;
· закупорка желчевыводящих протоков печени.
1. Гемолитическая (надпеченочная желтуха)
Гемолитическая желтуха развивается вследствие интенсивного гемолиза эритроцитов при гемолитических анемиях, вызванных сепсисом, лучевой болезнью, переливанием несовместимых групп крови, отравлением сульфаниламидами и т. д. Усиленный гемолиз эритроцитов приводит к интенсивному образованию в клетках РЭС непрямого билирубина. Печень не способна утилизировать в короткое время весь образующийся непрямой билирубин, он накапливается в крови и тканях. Так как печень обезвреживает повышенное количество непрямого билирубина, в больших количествах образуется прямой билирубин в печени.
2. Паренхиматозная (печёночная) желтуха.
Паренхиматозная желтуха обусловлена повреждением гепатоцитов при острых вирусных инфекциях, хроническом и токсическом гепатитах. Причина повышения концентрации билирубина – нарушение функций и некроз части печеночных клеток.
Биохимические нарушения.
1. В результате некроза гепатоцитов прямой билирубин частично попадает в кровь, его концентрация увеличивается. Прямой билирубин хорошо растворим в воде и экскретируется с мочой.
2. Экскреция жёлчи нарушена, в кишечник попадает меньше билирубина чем в норме, поэтому количество образующегося в толстом кишечнике стеркобилиногена также снижена. Кал гипохоличный.
3. При прогрессирующем гепатите нарушается процессы коньюгации билирубина в печени, вследствие этого в крови накапливается непрямой билирубин.
4. Нарушается процесс разрушения уробилиногена, поступающего в печень из кишечника по воротной вене. Он попадает в общий кровоток и экскретируется с мочой (в норме в моче отсутствует).
3. Механическая или обтурационная (подпеченочная) желтуха.
Механическая желтуха развивается при нарушении желчевыделения в двенадцатиперстную кишку. Основная причина – частичная или полная закупорка желчных протоков, например при жёлчно-каменной болезни, опухолях поджелудочной железы, желчного пузыря, печени. Так как нормальные пути экскреции коньюгированного билирубина заблокированы, происходит его поступление в кровь. В крови увеличивается содержание прямого билирубина, он выводится с мочой, придавая ей насыщенный оранжево-коричневый цвет. При полной закупорке общего желчного протока желчь не попадает в кишечник, не происходит образование стеркобилиногена, кал обесцвечен и в моче отсутствует уробилиноген.
4 вопрос. 1.Слюна здорового человека с адекватными зубными рядами имеет большие индивидуальные различия по концентрации и дебиту электролитов, общего белка, амилазы, тиреоидных и репродуктивных гормонов. Это свидетельствует о зависимости состава слюны от функционального состояния центра слюноотделения. Компоненты саливации положительно коррелированы с высокими коэффициентами по своим дебитам, то есть через напряжение саливации, и с низкими - по концентрации. Это свидетельствует о наличии генерализованных и селективных механизмов рефлекторного управления слюноотделением. Прямо зависимы компоненты саливации правой и левой слюнных желез.
2.Напряжение саливации, стимулированной раздражением вкусовых рецепторов натощак и после завтрака разное, одинаков состав электролитов, рН слюны, но постпрандиально выделяющаяся слюна имеет более высокую концентрацию белка, амилолитическую активность и дебиты основных компонентов слюны
3.У праворуких людей с интактными зубными рядами саливация по большинству параметров выше у правой, чем у левой околоушной железы. Дебиты компонентов слюны в секреции левых желез более вариабельны и коррелированны между собой с более высоким коэффициентом, чем в секреции правых желез.
4.Латерализованность саливации нарушается при неполных зубных рядах, так как парциальная односторонняя адентия трансформирует саливацию, увеличивая ее по большинству компонентов слюнной железы на стороне адентии, особенно по концентрации и дебиту Са2+.
5.Через год после протезирования неполных зубных рядов саливация у желез интактной и протезированной сторон восстанавливается по всем параметрам.