

Б.6 (1) Нервный центр и его свойства
Нервный центр - это совокупность нейронов, необходимых для осуществления определенного рефлекса или регуляции определенной функции. Любой нервный центр состоит из ядра и периферии. Ядерная часть нервного центра представляет собой функциональное объединение нейронов, в которое поступает основная информация от афферентных путей. Повреждение этого участка нервного центра приводит к повреждению или существенному нарушению осуществления данной функции. Периферическая часть нервного центра получает небольшую порцию афферентной информации, и её повреждение вызывает ограничение или уменьшение объёма выполняемой функции.
Рабочий отдел нервного центра - это отдел, ответственный за осуществление данной функции. Например, рабочий отдел дыхательного центра представлен центрами вдоха, выдоха и пневпотаксиса, расположенными в продолговатом мозге и варолиевом мосту, нарушение этого отдела вызывает остановку дыхания.
Регуляторный отдел нервного центра - это центр, расположенный в коре больших полушарий мозга и регулирующий активность рабочего отдела нервного центра.
В свою очередь, активность регуляторного отдела нервного центра зависит от состояния рабочего отдела, который получает афферентную информацию, и от внешних стимулов среды. Так, регуляторный отдел дыхательного центра расположен в лобной доле коры больших полушарий и позволяет произвольно регулировать лёгочную вентиляцию (глубину и частоту дыхания). Однако эта произвольная регуляция небезгранична и зависит от функциональной активности рабочего отдела, афферентной импульсации, отражающей состояние внутренней среды (в данном случае рН крови, концентрации углекислого газа и кислорода в крови).
Исполнительный отдел нервного центра - это двигательный центр, расположенный в спинном мозге и передающий информацию от рабочего отдела нервного центра к рабочим органам. Исполнительный отдел дыхательного нервного центра расположен в передних рогах грудного отдела спинного мозга и транслирует приказы рабочего центра к дыхательным мышцам.
С другой стороны, одни и те же нейроны головного и спинного мозга могут участвовать в регуляции разных функций. Например, клетки центра глотания участвуют в регуляции не только акта глотания, но и акта рвоты. Этот центр обеспечивает все последовательные стадии акта глотания: движение мышц языка, сокращение мышц мягкого нёба и его поднятия, последующее сокращение мышц глотки и пищевода при прохождении пищевого комка. Эти же нервные клетки обеспечиваю сокращение мышц мягкого нёба и его поднятие во время акта рвоты. Следовательно, одни и те же нервные клетки входят и в центр глотания, и в центр рвоты.
Свойства нервных центров зависят от их строения и механизмов передачи возбуждения в синапсах. Выделяются следующие свойства нервных центров.
1. Одностороннее проведение возбуждение в нервном центре. Возбуждение в ЦНС проводится в одном направлении с аксона на дендрит или тело клетки следующего нейрона. В основе этого свойства лежат особенности морфологической связи между нейронами.
Одностороннее проведение возбуждения -зависит от строения синапса и гуморальной природы передачи в нём импульса: медиатор, осуществляющий передачу возбуждения, выделяется только в пресинаптическом окончании, а рецепторы, воспринимающие медиатор, расположены на постсинаптической мембране.
2.Последействие - это способность сохранять возбуждение после окончания действия раздражителя, т.е. афферентных импульсов нет, а эфферентные продолжают действовать ещё некоторое время.
Последействие объясняется наличием следовой деполяризации. Если следовая деполяризация длительна, то на её фоне в течение нескольких миллисекунд могут возникать потенциалы действия (ритмическая активность нейрона), вследствие чего сохраняется ответная реакция. Но это дает сравнительно короткий эффект последействия. Более длительное последействие связано с наличием кольцевых связей между нейронами. В них возбуждение как бы само себя поддерживает, возвращаясь по коллатералям аксонам к первоначально возбужденному нейрону (рис. 3.9).
С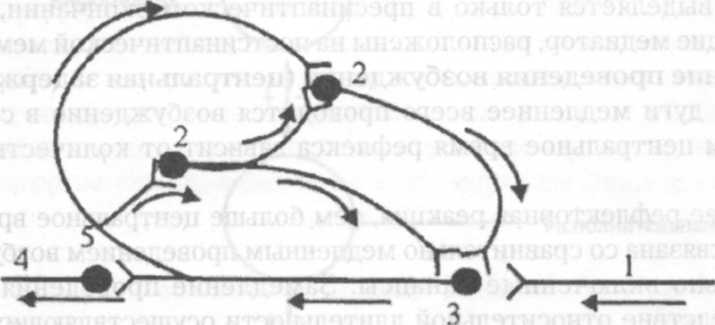 уммация.
Суммация
впервые была описана И.М. Сеченовым в
1863 г. и заключается в том, что слабые по
силе раздражители не вызывающие видимой
реакции, при частом повторении могут
суммироваться, создавать надпороговую
силу и вызывать эффект возбуждения.
Различают два вида суммации: последовательную
и пространственную.
уммация.
Суммация
впервые была описана И.М. Сеченовым в
1863 г. и заключается в том, что слабые по
силе раздражители не вызывающие видимой
реакции, при частом повторении могут
суммироваться, создавать надпороговую
силу и вызывать эффект возбуждения.
Различают два вида суммации: последовательную
и пространственную.
Последовательная суммация в синапсах возникает в том случае, когда по одному и тому же афферентному пути к центрам поступает несколько подпорогрвых импульсов. В результате суммации местного возбуждения, вызванного каждым подпороговым стимулом, возникает ответная реакция.
Пространственная суммация заключается в появлении рефлекторной реакции в ответ на два или несколько подпороговых стимулов, приходящих в нервный центр по разным афферентным путям (рис. 3.11
 Рис.
3.11. Свойство
нервного центра - суммация временная
(А) и последов-ная
(Б)
Рис.
3.11. Свойство
нервного центра - суммация временная
(А) и последов-ная
(Б)
Пространственную суммацию, как и последовательную, можно объяснить тем, что при подпороговом раздражении, пришедшем по одному афферентному пути, выделяется недостаточное количество медиатора для того, чтобы вызвать деполяризацию мембраны до критического уровня. Если же импульсы приходят одновременно несколькими афферентными путями к одному и тому же нейрону, в синапсах выделяется достаточное количество медиатора, необходимое для пороговой деполяризации и возникновения потенциала действия.
Окклюзия и облегчение. В зависимости от взаимного расположения ядерных и периферических зон разных нервных центров может проявиться при взаимодействии рефлексов явление окклюзии (закупорки) или облегчения (суммации).
Если происходит перекрывание ядер 2 нервных центров, то при раздражении афферентного поля первого нервного центра возникает условно 2 двигательных ответа. При активации только 2 центра, также возникает 2 двигательных ответа. При одновременной стимуляции 2 центров, двигательный ответ равен 3 единицам. Т.к. один и тот же мотонейрон относится одновременно к обоим нервным центрам.
При перекрывании периферических отделов разных нервных центров, при раздражении 1 центра – 2 ответа, то же самое при раздражении 2 центра. При одновременной возбуждении 2 центров, двигательный ответ равен 3 единицам. Т.к. мотонейроны, находящиеся в зоне перекрывания и не дающие ответа при изолированном раздражении нервных центров, получают при одновременной стимуляции обоих центров суммарную дозу медиатора, что приводит к пороговому уровню деполяризации.
Б.6 (2) Процесс свёртывания крови. При повреждении крупных сосудов (артерий, вен, артериол), также происходит образование тромбоцитарной пробки, но она не способна остановить кровотечение, так как легко вымывается током крови. Основное значение в этом процессе принадлежит свёртыванию крови, сопровождающемуся в конечном итоге образованием плотного фибринового сгустка.
Свёртыванием крови называют процесс образования сгустка - тромба - крови, состоящего из плазмы и форменных элементов. В основе свёртывания лежит образование нитей фибрина из белка плазмы фибриногена. Способность крови к свёртыванию является необходимой защитной реакцией, предохраняющей организм от кровопотерь. В плазме здорового человека имеются все компоненты, необходимые для её осуществления. Они формируют свёртывающую систему крови.
В настоящее время известно большое число соединений, относящихся к свёртывающей системе. Плазменные факторы свёртывания крови обозначают римскими цифрами, а тромбоцитарные - арабскими.
Плазменные факторы свёртывания - это различные компоненты плазмы, участвующие в образовании сгустка крови. Выделяют тринадцать таких факторов.
I- фибриноген - представляет собой самый крупномолекулярный белок плазмы, образуется в печени, его концентрация в крови составляет 200-400 мг%. При свёртывании крови фибриноген из состояния золя переходит в гель - фибрин, образующий основу кровяного сгустка. Содержание фибриногена резко возрастает при беременности, в послеоперационном периоде, при всех воспалительных процессах и инфекционных заболеваниях. Во время менструации, а также при болезнях печени его концентрация уменьшается. Кроме участия в свёртывании, фибрин служит структурным материалом для заживления ран.
- протромбин - является глюкопротеидом, образуется клетками печени при участии витамина К. Его концентрация в плазме составляет 2-4 мг%, переходящий в тромбин.
- тканевый тромбопластин - по своей природе представляет собой фосфолипид и входит в состав мембран всех клеток организма, в том числе эндотелия сосудов. Он необходим для образования тканевой протромбиназы.
- кальций - содержится в крови наполовину в виде ионов и наполовину в виде комплексов с белками плазмы. В свёртывании крови участвуют лишь ионы Са2+, которые необходимы для всех фаз свёртывания крови. Кровь доноров предохраняют от свёртывания путём связывания ионов Са2+ различными стабилизаторами (например, цитратом натрия).
V-VI - акцелератор-глобулин - белок, который образуется в печени и активизируется тромбином. Создает оптимальные условия для взаимодействия факторов X нП.
VII- конвертин - синтезируется в печени при участие витамина К, требуется для образования тканевой протромбиназы.
VIII- антигемофильный глобулин А - необходим для формирования протромбиназы. Его генетический дефицит служит причиной гемофилии А, протекающей с тяжёлыми кровотечениями.
IХ-фактор Кристмаса, или антигемофильный глобулин В - образуется в печени в присутствии витамина К. Его генетический дефицит вызывает гемофилию В.
Х-фактор Стюарта-Прауэра -Синтезируется в печени при участии витамина К, участвует в формировании и входит в состав тканевой и кровяной протромбиназ.
XI - плазменный предшественник тромбопластина - образуется в присутствии витамина К в печени, требуется для образования кровяной протромбиназы, где он активирует фактор IХ. Дефицит этого фактора служит причиной гемофилии С.
XII- фактор Хагемана - активируется при контакте с чужеродной поверхностью (например, местом повреждения сосуда), поэтому его называют также контактным фактором. Он является инициатором образования кровяной протромбиназы и всего процесса свёртывания крови. После активации он остаётся на поверхности повреждённого сосуда, что предупреждает генерализацию свёртывания крови. Генетический дефицит этого фактора служит причиной болезни Хагемана.
XIII- фибринстабилизирующий фактор (фибриназа) - содержится в плазме, клетках крови и тканях. По химической структуре он является гликопротеидом, синтезируется в печени и при свёртывании крови полностью потребляется. Этот фактор необходим для образования окончательного или нерастворимого фибрина. При врождённом дефиците фибриназы резко ухудшается заживление бытовых и хирургических ран, что говорит о необходимости этого фактора для регенерации.
Механизм свёртывания крови. Процесс свёртывания крови представляет собой преимущественно проферментно-ферментный каскад, в котором проферменты, переходят в активное состояние, приобретают способность активировать другие-факторы свёртывания крови. Процесс свёртывания крови можно разделить на несколько фаз.
1. Первая фаза - образование протромбиназы - является самой сложной и продолжительной. В этом процессе различают внешнюю (тканевую) и внутреннюю (кровяную) систему. Внешний путь запускается тканевым тромбопластином, который выделяется из стенок повреждённого сосуда и окружающих тканей. Во внутренней системе фосфолипиды и другие факторы поставляются самой кровью. В первую фазу образуется тканевая, тромбоцитарная, и эритроцитарная протромбиназы. Последние две часто называются кровяной протромбиназой. Образование тканевой протромбиназы длится 5-10 с, а кровяной - 5-10 мин.
2.Вторая фаза - переход протромбина в тромбин. Появление протромбиназы знаменует начало второй фазы свёртывания крови - образование тромбина. По сравнению с первой фазой, этот процесс протекает практически мгновенно 2-5 с. Такая скорость обусловлена тем, что протромбиназа адсорбирует протромбин на своей поверхности превращая его в тромбин. Этот процесс требует ряда факторов, указанных на схеме.
3.Третья фаза - переход фибриногена в фибрин. Этот процесс протекает в три этапа. На 1-м этапе под влиянием тромбина из фибриногена образуется фибрин-мономер. На 2-м этапе под влиянием солей кальция наступает полимеризация фибрин-мономеров и образуется фибрин-полимер. На 3-м этапе при участии фактора XIII и фибриназы тканей, тромбоцитов и эритроцитов образуется окончательный нерастворимый фибрин. Фибриназа образует прочные пептидные связи между соседними молекулами фибрин-полимера, что цементирует фибрин, увеличивает его механическую прочность и устойчивость к фибринолизу. Образование фибрина завершает процесс образования кровяного тромба. 4. После образования фибринового сгустка начинается последняя фаза свёртывания крови, включающая два процесса - ретракцию и фибринолиз. Ретракция обеспечивает уплотнение и закрепление тромба в повреждённом сосуде. Она осуществляется лишь при достаточном количестве тромбоцитов за счёт их сократительного белка тромбостенина. При сокращении он сжимает сгусток до 25-50% первоначального объёма, что закрепляет его в сосуде более надёжно. Ретракция заканчивается в течение 2-3 ч после образования сгустка.
Одновременно с ретракцией, но с меньшей скоростью начинается фибринолиз -расщепление фибрина, составляющего основу тромба. Главная функция фибринолиза -восстановление просвета закупоренного сгустком сосуда. Ферментом, разрушающим фибрин, является плазмин (иногда его называют «фибринолизин»), который в циркулирующей крови находится в неактивном состоянии.
Противосвёртывающие механизмы. Циркулирующая кровь имеет всё необходимое для свёртывания, однако остаётся жидкой. Сохранение жидкого состояния крови - одного из важнейших параметров гомеостаза - главная функция системы гемокоагуляции. Свёртывание крови представляет вторичное, защитное приспособление, включающееся при повреждении сосудов. Система гемокоагуляции в естественных условиях поддерживает жидкое состояние крови и оптимальное состояние стенок сосудов.
Жидкое состояние крови сохраняется за счёт многих механизмов:
свёртыванию крови препятствует гладкая поверхность эндотелия сосудов, что препятствует активации одного из факторов свёртывания крови (фактор Хагемана) и скручиванию (аггрегация) тромбоцитов;
стенки сосудов и форменные элементы крови имеют отрицательный заряд, что отталкивает клетки крови от стенок сосудов;
свёртыванию мешает большая скорость течения крови;
в крови есть вещества, разрушающие тромбин - антитромбины;
наличие в крови гепарина, который образуется в базофилах и в тучных клетках соединительной ткани. Много гепарина содержится в печени, мышцах и лёгких.
Регуляция свёртывания крови. Ещё в начале XX в. В. Кеннон отметил, что при болевом раздражении, эмоциях страха и гнева, т.е. состояниях, протекающих с возбуждением симпатического отдела вегетативной нервной системы и выделением большого количества адреналина, свёртывание крови ускоряется. Это наблюдается при всех стрессовых воздействиях, ускоряющих гемокоагуляцию на 25-50% и более с 5-10 мин до 3-4 мин. Совершенно ясно, что такое укорочение времени свёртывание может быть достигнуто лишь за счёт самой продолжительной фазы гемокоагуляции -образования протромбиназы.
Ускорение свёртывания крови называется гиперкоагулемией, а замедление -гипокоагулемией.
Гиперкоагулемия достигается за счёт расходования факторов свёртывания, поэтому после прекращения действия раздражителя на организм она сменяется вторичной гипокоагулемией. Развитие гиперкоагулемии подготавливает организм к более быстрому образованию сгустка при угрозе травмы или другого повреждения.
Раздражение блуждающего нерва (или внутривенное введение ацетилхолина) приводит к выделению из стенок сосудов веществ, аналогичных тем, которые выделяются при действии адреналина. Таким образом, в процессе эволюции в системе гемокоагуляции сформировалась лишь одна защитно-приспособительная реакция -гиперкоагулемия, направленная на срочную остановку кровотечения. Идентичность сдвигов гемокоагуляции при возбуждении симпатического и парасимпатического отделов ещё раз подтверждает тот факт, что первичной гипокоагулемии не существует. Она всегда вторична и развивается после первичной гиперкоагулемии вследствие использования части факторов свёртывания крови.
У здоровых людей ускорение гемокоагуляции, как правило, вызывает вторичную стимуляцию фибринолиза, что обеспечивает расщепление избытка фибрина, который образуется в результате усиления латентного микросвёртывания крови. Активация фибринолиза наблюдается при физической работе, эмоциях, болевом раздражении.