

Секреция медиатора
Содержимое синаптических пузырьков может выбрасываться в синаптическую щель путем экзоцитоза. При опорожнении одного синаптического пузырька в синаптичекую щель выбрасывается порция (квант) медиатора, которая включает около 10000 молекул.
Для активации экзоцитоза необходимы ионы Са++ . В состоянии покоя уровень Са++ в синаптическом окончании низок и выделения медиатора практически не происходит. Приход в синаптическое окончание возбуждения приводит к деполяризации пресинаптической мембраны и открытию потенциалчувствительных Са++ -каналов. Ионы Са++ поступают в цитоплазму синаптического окончания (рис. 3, А,Б) и активируют опорожнение синаптических пузырьков в синаптическую щель
33Мионевральный (нервно-мышечный) синапс – образован аксоном мотонейрона и мышечной клеткой.
Нервный импульс возникает в тригерной зоне нейрона, по аксону направляется к иннервируемой мышце, достигает терминали аксона и при этом деполяризует пресинаптическую мембрану.
После этого открываются натриевые и кальциевые каналы, и ионы Ca из среды, окружающей синапс, входят внутрь терминали аксона. При этом процессе броуновское движение везикул упорядочивается по направления к пресинаптической мембране. Ионы Ca стимулируют движение везикул. Достигая пресинап-тическую мембрану, везикулы разрываются, и освобождается ацетилхолин (4 иона Ca высвобождают 1 квант ацетилхолина). Синаптическая щель заполнена жидкостью, которая по составу напоминает плазму крови, через нее происходит диффузия АХ с преси-наптической мембраны на постсинаптическую, но ее скорость очень мала. Кроме того, диффузия возможна еще и по фиброзным нитям, которые находятся в синаптической щели. После диффузии АХ начинает взаимодействовать с хеморецепторами (ХР) и холи-нэстеразой (ХЭ), которые находятся на постсинапти-ческой мембране.
Холинорецептор выполняет рецепторную функцию, а холинэстераза выполняет ферментативную функцию. На постсинаптической мембране они расположены следующим образом:
ХР—ХЭ—ХР—ХЭ—ХР—ХЭ.
ХР + АХ = МПКП – миниатюрные потенциалы концевой пластины.
Затем происходит суммация МПКП. В результате сум-мации образуется ВПСП – возбуждающий постсинап-тический потенциал. Постсинаптическая мембрана за счет ВПСП заряжается отрицательно, а на участке, где нет синапса (мышечного волокна), заряд положительный. Возникает разность потенциалов, образуется потенциал действия, который перемещается по проводящей системе мышечного волокна.
ХЭ + АХ = разрушение АХ до холина и уксусной кислоты.
В состоянии относительного физиологического покоя синапс находятся в фоновой биоэлектрической активности. Ее значение заключается в том, что она повышает готовность синапса к проведению нервного импульса тем самым значительно облегчает передачу нервного возбуждения по синапсу. В состоянии покоя 1–2 пузырька в терминале аксона могут случайно подойти к пресинаптической мембране, в результате чего вступят с ней в контакт. Везикула при контакте с пресинап-тической мембраной лопается, и ее содержимое в виде 1 кванта АХ поступает в синаптическую щель, попадая при этом на постсинаптическую мембрану, где будет образовываться МПКН.
34
Взаимодействие медиатора с рецепторами постсинаптической мембраны. Это взаимодействие заключается в избирательном изменении проницаемости ионоселективных каналов эффекторной клетки в области активных центров связывания с медиатором. Взаимодействие медиатора со своими рецепторами может вызвать возбуждение или торможение нейрона, сокращение мышечной клетки, образование и выделение гормонов секреторными клетками. В случае увеличения проницаемости натриевых и кальциевых каналов усиливается поступление Na и Ca в клетку с последующей деполяризацией мембраны, возникновением ПД и дальнейшей передачей нервного импульса. Такие синапсы называются возбуждающими. Если повышается проницаемость калиевых каналов и каналов для хлора, наблюдается избыточный выход К из клетки с одновременной диффузией в нее Cl , что приводит к гиперполяризации мембраны, снижению ее возбудимости и развитию тормозных постсинаптических потенциалов. Передача нервных импульсов затрудняется или совсем прекращается. Такие синапсы называются тормозными.
Рецепторы, взаимодействующие с АХ, называются холинорецепторами. В функциональном отношении они разделяются на две группы: М - и Н-холинорецепторы. В синапсах скелетных мышц присутствуют только Н-холинорецепторы, тогда как в мышцах внутренних органов – преимущественно М-холинорецепторы.
Рецепторы, взаимодействующие с НА, называются адренорецепторами. В функциональном отношении они делятся на альфа- и бета-адренорецепторы. В постсинаптической мембране гладкомышечных клеток внутренних органов и кровеносных сосудов часто соседствуют оба вида адренорецепторов. Действие НА является деполяризующим, если он взаимодействует с альфа-адренорецепторами (сокращение мышечной оболочки стенок кровеносных сосудов или кишечника), или тормозным – при взаимодействии с бета-адренорецепторами (их расслабление).
Для синапсов с химической передачей возбуждения возникновение и развитие ВПСП связаны со следующим рядом последовательных явлений. В пресинаптическое окончание приходят ПД и вызывают их деполяризацию. При этом происходит мобилизация синаптических пузырьков и выделение квантов возбуждающего медиатора в синаптическую щель. Медиатор, взаимодействуя с хеморецепторным участком постсинаптической мембраны, увеличивает ее проницаемость в основном для ионов Na+. Усиленное движение Na+ через мембрану внутрь клетки приводит к возникновению электрического тока. В области синапса электрический ток направлен внутрь нейрона, а в прилежащих к синапсу участках — наружу. Электрический ток, направленный наружу, вызывает деполяризацию прилежащих к синапсу участков мембраны и является причиной возникновения ВПСП (рис. 11). Развитие ВПСП во времени характеризуется тем, что они возникают через некоторое время после прихода ПД в пресинаптическое окончание, так как в синапсах происходит синаптическая задержка. После этого в течение примерно 1-1,5 мс амплитуда ВПСП увеличивается (бремя подъема ВПСП), а затем наблюдается ее уменьшение, которое имеет экспоненциальный характер, и продолжается около 4-6 мс (время спада ВПСП). Характерным признаком ВПСП является их способность к суммации. Это означает, что амплитуда ВПСП, возникшего на приход в пресинаптическое окончание, например, одного импульса будет меньше по сравнению с амплитудой ВПСП, который разовьется на приход к этому же синапсу нескольких импульсов, следующих с меньшим временным интервалом, чем продолжительность ВПСП. Такой вид суммации называется «временной суммацией». Кроме временной имеется и пространственная суммация ВПСП. При этом амплитуда ВПСП увеличивается, если активируется не один, а несколько близкорасположенных возбуждающих синапсов. Возникновение ПД на основе ВПСП. Амплитуда ВПСП для большинства нейронов не превышает 10-20 мв, так как постсинаптическая деполяризация, достигнув определенного критического (порогового) уровня, становится причиной качественно нового процесса, в результате которого развиваются ПД. Рис. 13. Механиз¬мы генерации тормозного постсинаптического потенциала (ТПСП). А — синапс и направление электрического тока при его возбуждении; Б — амплитудные и временные параметры ТПСП (1) и электрического тока (2). Ионные механизмы возникновения и развития ПД в нейронах имеют некоторые характерные особенности. Эти особенности связаны с неодинаковой способностью различных участков мембраны тела и дендритов нейрона трансформировать ВПСП в потенциалы действия. Экспериментально установлено, что КУД начального сегмента аксона примерно в два раза ниже, чем у тела нейрона. Это означает, что наиболее возбудимой областью нервной клетки являются место отхождения аксона и его начальный немиелинизированный участок. Эта зона в смысле преобразования ВПСП в ПД наиболее эффективна и определяется как триггерная зона нейрона. Мембранный электрогенез сомы нейрона и крупных дендритов принципиально сходен с начальным сегментом аксона. Но возникновение распространяющихся ПД происходит при более высоком уровне деполяризации. Это подтверждается, в частности, тем, что ПД нейрона, возникающий при его антидромной или моносинаптической стимуляции и регистрируемый внутриклеточным электродом, имеет два компонента. В начале регистрируется потенциал начального сегмента, или НС-потенциал, а затем потенциал сомы нейрона, или СД-потенциал (рис. 12). Можно предположить, что дендриты сами по себе не способны генерировать ПД, а возникающие ВПСП, в силу их быстрого затухания, распространяются лишь на очень небольшое расстояние, часто не достигая тела нейрона. Однако развитие множественных ВПСП в зоне апикальных дендритов способно злектротонически изменять возбудимость нейронов и модулировать их импульсную активность. Механизмы генерации ТПСП. В пресинаптическом окончании тормозного синапса, активированного пришедшими сюда ПД, происходит выделение тормозного медиатора. Этот медиатор, проникнув в синаптическую щель, взаимодействует с хеморецепторными участками постсинаптической мембраны и увеличивает ее проницаемость преимущественно для ионов С1— и К+. Возникающий при этом электрический ток в области синапса направлен наружу, а в прилежащих к синапсу участках мембраны внутрь нейрона. Это приводит к гиперполяризации постсинаптической мембраны и возникновению ТПСП (рис. 13). ТПСП не являются зеркальным отображением ВПСП так как они имеют более продолжительный латентный период (по крайней мере, на время одной синаптической задержки) и меньшее время (около 3 мс) спада.
35
Существуют следующие методы исследования функций ЦНС:
1. Метод перерезок ствола мозга на различных уровнях. Например, между продолговатым и спинным мозгом.
2. Метод экстирпации (удаления) или разрушения участков мозга.
3. Метод раздражения различных отделов и центров мозга.
4. Анатомо-клинический метод. Клинические наблюдения за изменениями функций ЦНС при поражении ее каких-либо отделов с последующим патологоанатомическим исследованием.
5. Электрофизиологические методы:
а. электроэнцефалография – регистрация биопотенциалов мозга с поверхности кожи черепа. Методика разработана и внедрена в клинику Г. Бергером.
б. регистрация биопотенциалов различных нервных центров; используется вместе со стереотаксической техникой, при которой электроды с помощью микроманипуляторов вводят в строго определенное ядро.
в. метод вызванных потенциалов, регистрация электрической активности участков мозга при электрическом раздражении периферических рецепторов или других участков;
6. метод внутримозгового введения веществ с помощью микроинофореза;
7. хронорефлексометрия – определение времени рефлексов.
Электроэнцефалография (ЭЭГ) (электро- + др.-греч. ενκεφαλος — «головной мозг» + γραφω — «пишу», изображать) — раздел электрофизиологии, изучающий закономерности суммарной электрической активности мозга, отводимой с поверхности кожи головы, а также метод записи таких потенциалов (формирования электроэнцефалограмм). Также ЭЭГ — неинвазивный метод исследования функционального состояния головного мозга путем регистрации его биоэлектрической активности.
ЭЭГ — чувствительный метод исследования, он отражает малейшие изменения функции коры головного мозга и глубинных мозговых структур, обеспечивая миллисекундное временное разрешение, не доступное другим методам исследования мозговой активности, в частности ПЭТ и фМРТ.
Электроэнцефалография дает возможность качественного и количественного анализа функционального состояния головного мозга и его реакций при действии раздражителей. Запись ЭЭГ широко применяется в диагностической и лечебной работе (особенно часто при эпилепсии), в анестезиологии, а также при изучении деятельности мозга, связанной с реализацией таких функций, как восприятие, память, адаптация и т. д.
Выделяют три группы ЭЭГ: нормальные; пограничные между нормой и патологией; патологические.
Нормальными называются ЭЭГ, содержащие α- или β-ритмы, которые по амплитуде не превышают соответственно 100 и 15 мкВ в зонах их физиологической максимальной выраженности. На нормальной ЭЭГ взрослого бодрствующего человека могут наблюдаться Δ- и Θ-волны, по амплитуде не превышающие основной ритм, не носящие характера билатерально синхронных организованных разрядов или четкой локальности и охватывающие не более 15 % общего времени записи.
Пограничными называют ЭЭГ, выходящие за указанные рамки, но не имеющие характера явной патологической активности. К пограничным можно отнести ЭЭГ, на которых наблюдаются следующие феномены:
α-ритм с амплитудой, выше 100 мкВ, но ниже 150 мкВ, имеющий нормальное распределение, которое дает нормальные веретенообразные модуляции во времени;
β-ритм с амплитудой выше 15 мкВ, но ниже 40 мкВ, регистрирующийся в пределах отведения;
Δ- и Θ-волны, не превышающие по амплитуде доминирующего α-ритма и 50 мкВ, в количестве более 15 %, но менее 25 % общего времени регистрации, не имеющие характера билатерально-синхронных вспышек или регулярных локальных изменений;
четко очерченные вспышки α-волн амплитудой свыше 50 мкВ или β-волн амплитудой в пределах 20-30 мкВ на фоне плоской или низкоамплитудной активности;
а-волны заостренной формы в составе нормального α-ритма;
билатерально-синхронные генерализированные Δ- и Θ-волны с амплитудой до 120 мкВ при гипервентиляции.
36
Спинномозговая жидкость (liquor cerebro-spinalis) (СМЖ), наполняющая подпаутинные пространства головного и спинного мозга и мозговые желудочки, резко отличается от других жидкостей организма. С ней сходны только эндо- и перилимфа внутреннего уха, а также водянистая влага глаза. Выработка 70—90% спинномозговой жидкости осуществляется сосудистыми сплетениями III и IV желудочков, а также частью стенок боковых желудочков. 10—30% СМЖ вырабатывается тканями ЦНС и выделяется эпендимой вне области сосудистых сплетений. Сосудистые сплетения образованы ветвящимися выпячиваниями мягкой мозговой оболочки и покрыты кубической формы хориоидными эпендимоцитами [3]. Избирательная ультрафильтрация компонентов плазмы крови с образованием СМЖ происходит из капилляров в просвет желудочков через гема-то-ликворный барьер. Установлено, что клетки эпендимы способны также секретировать некоторые белки в СМЖ и частично поглощать вещества из СМЖ, очищая ее от продуктов метаболизма мозга.
Спинномозговая жидкость прозрачна, почти не содержит клеток (0—5 эритроцитов и 0—3 лейкоцитов в мм3). Установлено, что вода и соли СМЖ секретируются и резорбируются практически всей поверхностью в пределах суб-арахноидального пространства. Большинство компонетов СМЖ секретируются сплетениями сосудистой оболочки боковых желудочков, хотя некоторые из них также секретируются сплетением сосудистой оболочки третьего и четвертого желудочков. Объем спинномозговой жидкости равен 125—150 мл. В сутки ее образуется 400—500 мл. Время обновления половины объема СМЖ равняется трем часам. Основной ток СМЖ идет в каудальном направлении к отверстиям четвертого желудочка. СМЖ протекает через межжелудочковое отверстие Монро в третий желудочек, а затем через силь-виев водопровод среднего мозга в четвертый желудочек. Жидкость проходит через срединные и боковые апертуры в субарахноидальную цистерну. В субарахноидальном пространстве жидкость всасывается свободно на поверхности всех структур центральной нервной системы.
Хотя частичное всасывание СМЖ через клетки эпендимы происходит в самой системе желудочков, главным образом оно осуществляется уже после того, как СМЖ покинет эту систему через отверстие Люшка.
Спинномозговая жидкость выполняет многочисленные функции. Основными из них являются поддержание нормального гомеостаза нейронов и глии мозга, участие в их метаболизме (удаление метаболитов), механическое предохранение головного мозга. СМЖ образует гидростатическую оболочку вокруг мозга и его нервных корешков и сосудов, которые свободно взвешены в жидкости. Благодаря этому уменьшается натяжение нервов и сосудов. СМЖ обладает и интегративной функцией, благодаря переносу гормонов и других биологически активных веществ.
При накоплении избыточных количеств СМЖ развивается состояние, называемое гидроцефалией. Причиной этого может быть слишком интенсивное образование СМЖ в желудочках или чаще патологический процесс, создающий препятствие нормальному току СМЖ и ее выходу из полостей желудочков в субарах-ноидальное пространство, что может происходить при воспалительных процессах, сопровождающихся закупоркой отверстий Люшка или облитерацией третьего желудочка. Другой причиной этого может явиться атрезия, или закупорка водопровода.
При этом развиваются разнообразные симптомы поражения как головного мозга, так и глазного яблока. Так, при врожденном или приобретенном стенозе сильвиевого водопровода увеличивается третий желудочек, вызывая нарушения как сенсорных, так и двигательных функций глаза. Это может быть битемпораль-ная гемианопсия, нарушение взора кверху, нистагм и нарушение зрачкового рефлекса. Увеличение внутричерепного давления часто приводит к отеку диска зрительного нерва и позже ведет к атрофии зрительного нерва. Точный механизм этого явления пока полностью не понят. Предполагают, что повышение давления СМЖ в субарахноидальном простаранстве мозга приводит к увеличению внутричерепного давления и давления в субарахноидальном пространстве зрительного нерва. При этом сдавливаются вены и нарушается отток венозной крови.
37
труктурно-функциональной единицей нервной системы является нервная клетка, или нейрон, или нейроцит. Всю нервную систему можно представить как взаимосвязанную и взаимодействующую сеть из нескольких триллионов нервных клеток. Несмотря на их громадное разнообразие, можно говорить о ряде общих структурных и функциональных признаков, присущих всем нервным клеткам (рис. 5).
В нейроне выделяют следующие основные части: тело, отростки и их окончания.
Тело нейрона, размеры которого колеблются от 4 до 130 мкм, представляет собой скопление клеточной плазмы, в которой располагается ядро – носитель генетической информации, митохондрии – универсальные «генераторы» энергии, необходимой для обеспечения деятельности клетки, и большое количество структур, выполняющих различные специфические функции.
Поверхность нейрона, его оболочка, часто именуемая просто мембраной, не только обеспечивает обмен с окружающей средой, но, обладая свойствами полупроницаемой мембраны, является структурой, где развиваются сложные процессы биоэлектрогенеза, лежащие в основе главных функций нервной клетки.
Отростки нервных клеток являются выростами цитоплазмы. Различают два вида отростков. Дендриты – короткие, древовидно ветвящиеся, постепенно истончаются и заканчиваются в окружающих тканях. Количество их достигает десяти, они многократно увеличивают поверхность клетки.
Помимо дендритов нервная клетка всегда имеет один аксон (или нейрит). Этот отросток всегда более крупный, длинный (до 1 м) и менее ветвистый. Аксон заканчивается синапсом, при помощи которого он функционально взаимодействует с иннервируемыми структурами.
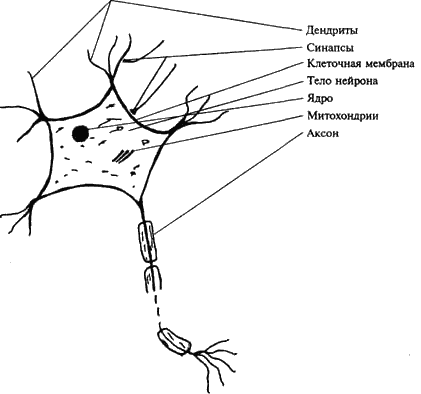
Рис. 5. Строение нервной клетки (упрощенная схема)
По своей функциональной значимости в составе рефлекторной дуги различают три вида нейронов:
рецепторные (чувствительные, афферентные), имеющие чувствительные нервные окончания, которые способны воспринимать раздражения из внешней или внутренней среды;
эффекторные (эфферентные), окончания аксонов которых передают нервный сигнал на рабочий орган;
ассоциативные (вставочные, центральные), являющиеся промежуточными в составе рефлекторной дуги и передающие информацию с чувствительного нейрона на эффекторные.
Следует иметь в виду, что на теле и отростках большинства нервных клеток имеется очень большое количество синапсов, через которые поступает информация с других нейронов.
Несмотря на громадное морфологическое и функциональное разнообразие нейронов, можно выделить ряд ключевых свойств и функций.
К числу наиболее важных свойств относятся:
1. Наличие трансмембранной разности потенциалов, т. е. между наружной и внутренней поверхностями оболочки нейрона в покое регистрируется разность потенциала порядка 90 мВ, наружная поверхность электроположительна по отношению к внутренней. Величина и направление трансмембранного тока меняются в зависимости от состояния нейрона.
2. Очень высокая чувствительность к некоторым химическим веществам (медиаторам) и электрическому току.
3. Способность к нейросекреции, т. е. к синтезу и выделению в окружающую среду или в синаптическую щель биологически активных веществ.
4. Высокий уровень энергетических процессов, что обусловливает необходимость постоянного притока основного источника энергии – глюкозы и кислорода, необходимого для окисления.
Принято различать следующие функции нейрона:
1. Воспринимающая – эта функция представлена двумя механизмами. Во-первых, чувствительные окончания дендритов способны обеспечить рецепцию, т. е. трансформацию специфической энергии раздражителя внешней или внутренней среды в неспецифический процесс нервного возбуждения, нервный импульс, который по отростку распространяется по направлению к телу нервной клетки. Во-вторых, на всех частях нейрона имеются многочисленные (до нескольких десятков тысяч) синапсы, при помощи которых химическим путем возбуждение передается от одного нейрона к другому. Химические вещества, осуществляющие эту передачу, обозначают медиаторы (или нейротрансмиттеры). К их числу, в частности, относятся адреналин, норадреналин, дофамин, серотонин, ацетилхолин, гамма-аминомасляная кислота и многие другие. В результате воздействия медиатора в теле нервной клетки развивается возбуждение и возникновение нервного импульса или снижение возбудимости нейрона – его торможение.
2. Интегративная – обработка одновременно или в течение короткого интервала времени поступающих нервных сигналов по механизму их алгебраической суммации, в результате которой на выходе нейрона формируется сигнал, несущий в себе информацию всех суммированных сигналов.
3. Мнестическая, основанная на существовании тонких молекулярных биофизических процессов, сохраняющих след от всякого предыдущего воздействия и благодаря этому трансформирующих характер ответной реакции на всякое последующее. По существу, это элементарная форма памяти и научения.
4. Проводниковая функция, суть которой состоит в том, что от тела нейрона по аксону к его окончанию в естественных условиях только в одном этом направлении распространяется, не затухая, нервный импульс. Скорость его распространения в зависимости от морфофункциональных особенностей проводника колеблется от нескольких сантиметров до 100–120 метров в секунду.
5. Передающая, проявляющаяся в том, что нервный импульс, достигнув окончания аксона, который, собственно, уже входит в структуру синапса, обусловливает выделение медиатора – непосредственного передатчика возбуждения к другому нейрону или исполнительному органу.
Часто в бытовых разговорах приходится слышать сожалеющее высказывание, что нервные клетки не восстанавливаются. Да, применительно к телу нейрона, это действительно так, и в ряде случаев это действительно плохо. Но следует также иметь в виду, что количество нейронов у человека значительно превышает его потребности на протяжении всей жизни. И, кроме того, как указывалось выше, нервные клетки на протяжении жизни человека «обучаются», «приобретают опыт», а потому включение в слаженный нейрональный ансамбль «необученного» элемента затруднило бы его работу.
38
ФИЗИОЛОГИЯ СИНАПСОВ
Синапсами называются контакты, которые устанавливают нейроны как самостоятельные образования. Синапс представляет собой сложную структуру и состоит из пресинаптической части (окончание аксона, передающее сигнал), синаптической щели и постсинаптической части (структура воспринимающей клетки).
Классификация синапсов. Синапсы классифицируются по местоположению, характеру действия, способу передачи сигнала.
По местоположению выделяют нервно-мышечные синапсы и нейронейрональные, последние в свою очередь делятся на аксосоматические, аксоаксональные, аксодендритические, дендросоматические.
По характеру действия на воспринимающую структуру синапсы могут быть возбуждающими и тормозящими.
По способу передачи сигнала синапсы делятся на электрические, химические, смешанные.
Характер взаимодействия нейронов. Определяется способом этого взаимодействия: дистантное, смежное, контактное.
Дистантное взаимодействие может быть обеспечено двумя нейронами, расположенными в разных структурах организма. Например, в клетках ряда структур мозга образуются нейрогормоны, нейропептиды, которые способны воздействовать гуморально на нейроны других отделов.
Смежное взаимодействие нейронов осуществляется в случае, когда мембраны нейронов разделены только межклеточным пространством. Обычно такое взаимодействие имеется там, где между мембранами нейронов нет глиальных клеток. Такая смежность характерна для аксонов обонятельного нерва, параллельных волокон мозжечка и т. д. Считают, что смежное взаимодействие обеспечивает участие соседних нейронов в выполнении единой функции. Это происходит, в частности, потому, что метаболиты, продукты активности нейрона, попадая в межклеточное пространство, влияют на соседние нейроны. Смежное взаимодействие может в ряде случаев обеспечивать передачу электрической информации от нейрона к нейрону.
Контактное взаимодействие обусловлено специфическими контактами мембран нейронов, которые образуют так называемые электрические и химические синапсы.
Электрические синапсы. Морфологически представляют собой слияние, или сближение, участков мембран. В последнем случае синаптическая щель не сплошная, а прерывается мостиками полного контакта. Эти мостики образуют повторяющуюся ячеистую структуру синапса, причем ячейки ограничены участками сближенных мембран, расстояние между которыми в синапсах млекопитающих 0,15—0,20 нм. В участках слияния мембран находятся каналы, через которые клетки могут обмениваться некоторыми продуктами. Кроме описанных ячеистых синапсов, среди электрических синапсов различают другие — в форме сплошной щели; площадь каждого из них достигает 1000 мкм, как, например, между нейронами ресничного ганглия.
Электрические синапсы обладают односторонним проведением возбуждения. Это легко доказать при регистрировании электрического потенциала на синапсе: при раздражении афферентных путей мембрана синапса деполяризуется, а при раздражении эфферентных волокон — гиперполяризуется. Оказалось, что синапсы нейронов с одинаковой функцией обладают двусторонним проведением возбуждения (например, синапсы между двумя чувствительными клетками), а синапсы между разнофункциональными нейронами (сенсорные и моторные) обладают односторонним проведением. Функции электрических синапсов заключаются прежде всего в обеспечении срочных реакций организма. Этим, видимо, объясняется расположение их у животных в структурах, обеспечивающих реакцию бегства, спасения от опасности и т. д.
Электрический синапс сравнительно мало утомляем, устойчив к изменениям внешней и внутренней среды. Видимо, эти качества наряду с быстродействием обеспечивают высокую надежность его работы.
Химические синапсы. Структурно представлены пресинаптической частью, синаптической щелью и постсинаптической частью. Пресинаптическая часть химического синапса образуется расширением аксона по его ходу или окончания (рис. 2.19). В пресинаптической части имеются агранулярные и гранулярные пузырьки. Пузырьки (кванты) содержат медиатор. В пресинаптическом расширении находятся митохондрии, обеспечивающие синтез медиатора, гранулы гликогена и др. При многократном раздражении пресинаптического окончания запасы медиатора в синаптических пузырьках истощаются. Считают, что мелкие гранулярные пузырьки содержат норадреналин, крупные — другие катехоламины. Агранулярные пузырьки содержат ацетилхолин. Медиаторами возбуждения могут быть также производные глутаминовой и аспарагиновой кислот.
Синаптические контакты могут быть между аксоном и дендритом (аксодендритические), аксоном и сомой клетки (аксосоматические), аксонами (аксоаксональные), дендритами (дендродендритические), дендритами и сомой клетки.
Действие медиатора на постсинаптическую мембрану заключается в повышении ее проницаемости для ионов Na+. Возникновение потока ионов Na+ из синаптической щели через постсинаптическую мембрану ведет к ее деполяризации и вызывает генерацию возбуждающего постсинаптического потенциала (ВПСП) (см. рис. 2.19).
Для синапсов с химическим способом передачи возбуждения характерны синоптическая задержка проведения возбуждения, длящаяся около 0,5 мс, и развитие постсинаптического потенциала (ПСП) в ответ на пресинаптический импульс. Этот потенциал при возбуждении проявляется в деполяризации постсинаптической мембраны, а при торможении — в гиперполяризации ее, в результате чего развивается тормозной постсинаптический потенциал (ТПСП). При возбуждении проводимость постсинаптической мембраны увеличивается.
ВПСП возникает в нейронах при действии в синапсах ацетил холина, норадреналина, дофамина, серотонина, глутаминовой кислоты, вещества Р.
ТПСП возникает при действии в синапсах глицина, гамма-аминомасляной кислоты. ТПСП может развиваться и под действием медиаторов, вызывающих ВПСП, но в этих случаях медиатор вызывает переход постсинаптической мембраны в состояние гиперполяризации.
Для распространения возбуждения через химический синапс важно, что нервный импульс, идущий по пресинаптической части, полностью гасится в синаптической щели. Однако нервный импульс вызывает физиологические изменения в пресинаптической части мембраны. В результате у ее поверхности скапливаются синаптические пузырьки, изливающие медиатор в синаптическую щель.
Переход медиатора в синаптическую щель осуществляется путем экзоцитоза: пузырек с медиатором соприкасается и сливается с пресинаптической мембраной, затем открывается выход в синаптическую щель и в нее попадает медиатор. В покое медиатор попадает в синаптическую щель постоянно, но в малом количестве. Под влиянием пришедшего возбуждения количество медиатора резко возрастает. Затем медиатор перемещается к постсинаптической мембране, действует на специфические для него рецепторы и образует на мембране комплекс медиатор—рецептор. Данный комплекс изменяет проницаемость мембраны для ионов К+ и Na+, в результате чего изменяется ее потенциал покоя.
В зависимости от природы медиатора потенциал покоя мембраны может снижаться (деполяризация), что характерно для возбуждения, или повышаться (гиперполяризация), что типично для торможения. Величина ВПСП зависит от количества выделившегося медиатора и может составлять 0,12—5,0 мВ. Под влиянием ВПСП деполяризуются соседние с синапсом участки мембраны, затем деполяризация достигает аксонного холмика нейрона, где возникает возбуждение, распространяющееся на аксон.
В тормозных синапсах этот процесс развивается следующим образом: аксонное окончание синапса деполяризуется, что приводит к появлению слабых электрических токов, вызывающих мобилизацию и выделение в синаптическую щель специфического тормозного медиатора. Он изменяет ионную проницаемость постсинаптической мембраны таким образом, что в ней открываются поры диаметром около 0,5 нм. Эти поры не пропускают ионы Na+ (что вызвало бы деполяризацию мембраны), но пропускают ионы К+ из клетки наружу, в результате чего происходит гиперполяризация постсинаптической мембраны.
Такое изменение потенциала мембраны вызывает развитие ТПСП. Его появление связывают с выделением в синаптическую щель специфического медиатора. В синапсах разных нервных структур роль тормозного медиатора могут выполнять различные вещества. В ганглиях моллюсков роль тормозного медиатора выполняет ацетилхолин, в ЦНС высших животных — гамма-аминомасляная кислота, глицин.
Нервно-мышечные синапсы обеспечивают проведение возбуждения с нервного волокна на мышечное благодаря медиатору ацетилхолину, который при возбуждении нервного окончания переходит в синаптическую щель и действует на концевую пластинку мышечного волокна. Следовательно, как и межнейронный синапс, нервно-мышечный синапс имеет пресинаптическую часть, принадлежащую нервному окончанию, синаптическую щель, постсинаптическую часть (концевая пластинка), принадлежащую мышечному волокну.
В пресинаптической терминали образуется и скапливается в виде пузырьков ацетилхолин. При возбуждении электрическим импульсом, идущим по аксону, пресинаптической части синапса ее мембрана становится проницаемой для ацетилхолина.
Эта проницаемость возможна благодаря тому, что в результате деполяризации пресинаптической мембраны открываются ее кальциевые каналы. Ион Са2+ входит в пресинаптическую часть синапса из синаптической щели. Ацетилхолин высвобождается и проникает в синаптическую щель. Здесь он взаимодействует со своими рецепторами постсинаптической мембраны, принадлежащей мышечному волокну. Рецепторы, возбуждаясь, открывают белковый канал, встроенный в липидный слой мембраны. Через открытый канал внутрь мышечной клетки проникают ионы Na+, что приводит к деполяризации мембраны мышечной клетки, в результате развивается так называемый потенциал концевой пластинки (ПКП). Он вызывает генерацию потенциала действия мышечного волокна.
Нервно-мышечный синапс передает возбуждение в одном направлении: от нервного окончания к постсинаптической мембране мышечного волокна, что обусловлено наличием химического звена в механизме нервно-мышечной передачи.
Скорость проведения возбуждения через синапс намного меньше, чем по нервному волокну, так как здесь тратится время на активацию пресинаптической мембраны, переход через нее кальция, выделение ацетилхолина в синаптическую щель, деполяризацию постсинаптической мембраны, развитие ПКП.
Синаптическая передача возбуждения имеет рад свойств:
1) наличие медиатора в пресинаптической части синапса;
2) относительная медиаторная специфичность синапса, т. е. каждый синапс имеет свой доминирующий медиатор;
3) переход постсинаптической мембраны под влиянием медиаторов в состояние де- или гиперполяризации;
4) возможность действия специфических блокирующих агентов на рецептирующие структуры постсинаптической мембраны;
5) увеличение длительности постсинаптического потенциала мембраны при подавлении действия ферментов, разрушающих синаптической медиатор;
6) развитие в постсинаптической мембране ПСП из миниатюрных потенциалов, обусловленных квантами медиатора;
7) зависимость длительности активной фазы действия медиатора в синапсе от свойств медиатора;
8) односторонность проведения возбуждения;
9) наличие хемочувствительных рецепторуправляемых каналов постсинаптической мембраны;
10) увеличение выделения квантов медиатора в синаптическую щель пропорционально частоте приходящих по аксону импульсов;
11) зависимость увеличения эффективности синаптической передачи от частоты использования синапса («эффект тренировки»);
12) утомляемость синапса, развивающаяся в результате длительного высокочастотного его стимулирования. В этом случае утомление может быть обусловлено истощением и несвоевременным синтезом медиатора в пресинаптической части синапса или глубокой, стойкой деполяризацией постсинаптической мембраны (пессимальное торможение).
Перечисленные свойства относятся к химическим синапсам. Электрические синапсы имеют некоторые особенности, а именно: малую задержку проведения возбуждения; возникновение деполяризации как в пре-, так и в постсинаптической частях синапса; наличие большей площади синаптической щели в электрическом синапсе, чем в химическом.
Синаптические медиаторы являются веществами, которые имеют специфические инактиваторы. Например, ацетилхолин инактивируется ацетилхолинэстеразой, норадреналин — моноаминоксидазой, катехолометилтрансферазой.
Неиспользованный медиатор и его фрагменты всасываются обратно в пресинаптическую часть синапса.
Ряд химических веществ крови и постсинаптической мембраны изменяет состояние синапса, делает его неактивным. Так, простагландины тормозят секрецию медиатора в синапсе. Другие вещества, называемые блокаторами хеморецепторных каналов, прекращают передачу в синапсах. Например, ботулинический токсин, марганец блокируют секрецию медиатора в нервно-мышечном синапсе, в тормозящих синапсах ЦНС. Тубокурарин, атропин, стрихнин, пенициллин, пикротоксин и др. блокируют рецепторы в синапсе, в результате чего медиатор, попав в синаптическую щель, не находит своего рецептора.
В то же время выделены вещества, которые блокируют системы, разрушающие медиаторы. К ним относят эзерин, фосфорорганические соединения.
В нервно-мышечном синапсе в норме ацетилхолин действует на синаптическую мембрану короткое время (1—2 мс), так как сразу же начинает разрушаться ацетилхолинэстеразой. В случаях, когда этого не происходит и ацетилхолин не разрушается на протяжении сотни миллисекунд, его действие на мембрану прекращается и мембрана не деполяризуется, а гиперполяризуется и возбуждение через этот синапс блокируется.
Блокада нервно-мышечной передачи может быть вызвана следующими способами:
1) действие местноанестезирующих веществ, которые блокируют возбуждение в пресинаптической части;
2) блокада высвобождения медиатора в пресинаптической части (например, ботулинический токсин);
3) нарушение синтеза медиатора, например при действии гемихолиния;
4) блокада рецепторов ацетилхолина, например при действии бунгаротоксина;
5) вытеснение ацетилхолина из рецепторов, например действие кураре;
6) инактивация постсинаптической мембраны сукцинилхолином, декаметонием и др.;
7) угнетение холинэстеразы, что приводит к длительному сохранению ацетилхолина и вызывает глубокую деполяризацию и инактивацию рецепторов синапсов. Такой эффект наблюдается при действии фосфорорганических соединений.
Специально для снижения тонуса мышц, особенно при операциях, используют блокаду нервно-мышечной передачи миорелаксантами; деполяризующие мышечные релаксанты действуют на рецепторы субсинаптической мембраны (сукцинилхолин и др.), недеполяризующие мышечные релаксанты, устраняющие действие ацетилхолина на мембрану по конкуренции (препараты группы кураре).
39
Медиаторами ЦНС являются многие химические вещества, разнородные в структурном отношении (в головном мозге обнаружено около 30 биологически активных веществ). По химическому строению их можно разделить на несколько групп, главными из которых являются моноамины, аминокислоты и полипептиды. Достаточно широко распространенным медиатором является ацетилхолин.
А. Ацетилхолин. Встречается в различных отделах ЦНС, известен в основном как возбуждающий медиатор: в частности, является медиатором α-мотонейронов спинного мозга, иннервирующих скелетную мускулатуру. С помощью ацетилхолина α -мотонейроны по коллатералям своих аксонов передают возбуждение на тормозные клетки Реншоу. В ретикулярной формации ствола мозга, в гипоталамусе обнаружены М- и N-холинорецепторы. При взаимодействии ацетилхолина с рецепторным белком последний изменяет свою конформацию, в результате чего открывается ионный канал. Тормозное влияние ацетилхолин оказывает с помощью М-холинорецепторов в глубоких слоях коры большого мозга, в стволе мозга, хвостатом ядре.
Б. Моноамины. Выделяют катехоламины, серотонин и гистамин. Большинство из них в значительных количествах содержится в нейронах ствола мозга, в меньших количествах они обнаруживаются в других отделах ЦНС.
Катехоламины обеспечивают возникновение процессов возбуждения и торможения, например, в промежуточном мозге, черной субстанции, лимбической системе, полосатом теле.
С помощью серотонина в нейронах ствола мозга передаются возбуждающие и тормозящие влияния, в коре мозга — тормозящие влияния. Серотонин содержится главным образом в структурах, имеющих отношение к регуляции вегетативных функций. Особенно много его в лимбической системе, ядрах шва. В нейронах названных структур выявлены ферменты, участвующие в синтезе серотонина. Аксоны этих нейронов проходят в бульбо-спинальных путях и оканчиваются на нейронах различных сегментов спинного мозга. Здесь они контактируют с клетками преганглионарных симпатических нейронов и со вставочными нейронами желатинозной субстанции. Полагают, что часть этих так называемых симпатических нейронов, а может быть и все, являются серотонинергическими нейронами вегетативной нервной системы. Их аксоны, согласно данным некоторых авторов, идут к органам пищеварительного тракта и стимулируют их сокращение.
Гистамин в довольно высокой концентрации обнаружен в гипофизе и срединном возвышении гипоталамуса. В остальных отделах ЦНС уровень гистамина очень низкий. Медиаторная роль его изучена мало. Выделяют Н1- и Н2-гистаминорецепторы. Н1-рецепторы имеются в гипоталамусе и участвуют в регуляции потребления пищи, терморегуляции, секреции пролактина и антидиуретического гормона. Н2-рецепторы обнаружены на глиальных клетках.
В. Аминокислоты. Кислые аминокислоты (глицин, γ-аминомасляная кислота) являются тормозными медиаторами в синапсах ЦНС и действуют на тормозные рецепторы (см. раздел 4.8). Нейтральные аминокислоты (α -глутамат, α -аспартат) передают возбуждающие влияния и действуют на соответствующие возбуждающие рецепторы. Предполагают, что глутамат может быть медиатором афферентов в спинном мозге. Рецепторы глутаминовой и аспарагиновой аминокислот имеются на клетках спинного мозга, мозжечка, таламуса, гиппокампа, коры большого мозга. Полагают, что глутамат - самый распространенный медиатор ЦНС.
Г. Полипептиды. В синапсах ЦНС они также выполняют медиаторную функцию. В частности, субстанция Р является медиатором нейронов, передающих сигналы боли. Особенно много этого полипептида в дорсальных корешках спинного мозга. Это послужило основанием к предположению, что субстанция Р может быть медиатором чувствительных нервных клеток в области их переключения на вставочные нейроны. Субстанция Р в больших количествах содержится в гипоталамической области. Различают два вида рецепторов субстанции Р: рецепторы типа SР-Р, расположенные на нейронах мозговой перегородки, и рецепторы типа SР-Е, расположенные на нейронах коры большого мозга.
Энкефалины и эндорфины - медиаторы нейронов, блокирующих болевую импульсацию. Они реализуют свое влияние посредством соответствующих опиатных рецепторов, которые особенно плотно располагаются на клетках лимбической системы; много их также на клетках черной субстанции, ядрах промежуточного мозга и солитарного тракта, имеются они на клетках голубого пятна, спинного мозга. Их лигандами являются [3-эндорфин, динорфин, лей- и мет-энкефалины.
Ангиотензин участвует в передаче информации о потребности организма в воде, люлиберин — в половой активности.
Физиологические эффекты действия некоторых медиаторов головного мозга. Норадреналин регулирует настроение, эмоциональные реакции, обеспечивает поддержание бодрствования, участвует в механизмах формирования некоторых фаз сна, сновидений; дофамин -в формировании чувства удовольствия, регуляции эмоциональных реакций, поддержании бодрствования. Дофамин полосатого тела регулирует сложные мышечные движения. Серотонин ускоряет процессы обучения, формирования болевых ощущений, сенсорное восприятие, засыпание, ангиотензин - повышение АД, торможение синтеза катехоламинов, стимулирует секрецию гормонов; информирует ЦНС об осмотическом давлении крови. Олигопептиды — медиаторы настроения, полового поведения; передачи ноцицептивного возбуждения от периферии в ЦНС, формирования болевых ощущений. Эндорфины, энкефалины, пептид, вызывающий дельта-сон, обусловливают антиболевые реакции, повышение устойчивости к стрессу, сон. Простагландины вызывают повышение свертываемости крови, изменение тонуса гладких мышц, усиление физиологического эффекта медиаторов и гормонов. Мозгоспецифичные белки различных отделов головного мозга влияют на процессы обучения.
Согласно принципу Дейла, один нейрон синтезирует и использует один и тот же медиатор или одни и те же медиаторы во всех разветвлениях своего аксона. Кроме основного медиатора, как выяснилось, в окончаниях аксона могут выделяться и другие — сопутствующие медиаторы, играющие модулирующую роль.
Эффект действия медиатора зависит в основном от свойств ионных каналов постсинаптической мембраны. Это явление особенно ярко демонстрируется при сравнении эффектов отдельных медиаторов в ЦНС и в периферических синапсах организма. Ацетилхолин, например, в коре мозга при микроаппликациях на разные нейроны может вызывать возбуждение и торможение, в синапсах сердца — торможение, в синапсах гладкой мускулатуры пищеварительного тракта — возбуждение. Катехоламины вызывают возбуждение и торможение в стволе мозга. 40
Процессы распространения возбуждения и торможения в ЦНС также имеют особенности. Рассматривая их в режиме распространения возбуждения в ЦНС, можно выделить следующие моменты.
1. Одностороннее проведения возбуждения. В ЦНС, ее центрах, внутри рефлекторных дуг и нейронных цепей возбуждение, как правило, идет в одном направлении: от афферентного нейрона к эфферентному, а не наоборот. Обусловлено это особенностями расположения и характером функционирования химического синапса. В целом это организует деятельность ЦНС и является одним из ее координационных принципов.
2. Суммация возбуждения (или торможения): Один импульс (даже, если он достаточно сильный) не в состоянии вызвать возбуждение на мембране синапса. Для этого необходима серия таких импульсов (по силе и частоте), которые могут достичь критического уровня поляризации мембраны, в результате чего начинает действовать механизм передачи этого возбуждения (пример с картинкой: калиево- натриевый насос). Если раздражение не достигает порогового (т.е. критического) уровня, то возбуждение не наступает.
3. Явление окклюзии (закупорки). Один и тот же нейрон может передавать сигналы на ряд других нейронов, в результате чего возникает определенный эффект. Например, нейрон активизировал 10 мышечных волокон в мышце, вследствие чего возникло ее определенное электрическое напряжение. Второй нейрон тоже возбуждает 10 других мышечных волокон. Но если оба нейрона возбуждать одновременно, то суммарная электрическая активность мышцы будет на 20% меньше, чем при последовательном возбуждении. Оказывается часть волокон у 1-го и 2-го нейрона были общими при возбуждении. Это и вызвало блокировку некоторой части информации и ее закупорку (т.е. явление окклюзии).
4. Трансформация ритма возбуждения (или торможения). В отличие от скелетной мышцы или аксона нейрон способен трансформировать ритм возбуждений, приходящий к нему. Например, на него поступает импульс с частотой 25 Гц, а нейрон в ответ на это, возбуждаясь, генерирует 50 Гц или уменьшает силу импульса с 25 до 15 Гц и т.д.
5. Последействие — один из вариантов этого свойства. Поступающий импульс может минутами или даже часами пробегать небольшой отрезок нейронной цепи. Это обеспечивает перевод «следа» из кратковременной в долговременную память.
6. Утомление нервных центров. При длительном возбуждении одного и того же нейрона в его синапсе снижается содержание медиатора, который влияет на работоспособность нейрона.
41
Свойства нервных центров
Рефлекторная деятельность организма во многом определяется общими свойствами нервных центров.
Нервный центр — совокупность структур центральной нервной системы, координированная деятельность которых обеспечивает регуляцию отдельных функций организма или определенный рефлекторный акт. Представление о структурно-функциональной основе нервного центра обусловлено историей развития учения о локализации функций в центральной нервной системе. На смену старым теориям об узкой локализации, или эквипотенциальности, высших отделов головного мозга, в частности коры большого мозга, пришло современное представление о динамической локализации функций, основанное на признании существования четко локализованных ядерных структур нервных центров и менее определенных рассеянных элементов анализаторных систем мозга. При этом с цефализацией нервной системы растут удельный вес и значимость рассеянных элементов нервного центра, внося существенные различия в анатомических и физиологических границах нервного центра. В результате функциональный нервный центр может быть локализован в разных анатомических структурах. Например, дыхательный центр представлен нервными клетками, расположенными в спинном, продолговатом, промежуточном мозге, в коре большого мозга.
Нервные центры имеют ряд общих свойств, что во многом определяется структурой и функцией синаптических образований.
1. Односторонность проведения возбуждения. В рефлекторной дуге, включающей нервные центры,
процесс возбуждения распространяется в одном направлении (от входа, афферентных путей к выходу, эфферентным путям).
2. Иррадиация возбуждения. Особенности структурной организации центральных нейронов, огромное
число межнейронных соединений в нервных центрах существенно модифицируют (изменяют) направление распространения процесса возбуждения в зависимости от силы раздражителя и функционального состояния центральных нейронов. Значительное увеличение силы раздражителя приводит к расширению области вовлекаемых в процесс возбуждения центральных нейронов — иррадиации возбуждения.
3. Суммация возбуждения. В работе нервных центров значительное место занимают процессы пространственной и временной суммации возбуждения, основным нервным субстратом которой является постсинаптическая мембрана. Процесс пространственной суммации афферентных потоков возбуждения облегчается наличием на мембране нервной клетки сотен и тысяч синаптических контактов. Процессы временной суммации обусловлены суммацией ВПСП на постсинаптической мембране.
4. Наличие синаптической задержки. Время рефлекторной реакции зависит в основном от двух факторов: скорости движения возбуждения по нервным проводникам и времени распространения возбуждения с одной клетки на другую через синапс. При относительно высокой скорости распространения импульса по нервному проводнику основное время рефлекса приходится на синаптическую передачу возбуждения (синаптическая задержка). В нервных клетках высших животных и человека одна синаптическая задержка примерно равна 1 мс. Если учесть, что в реальных рефлекторных дугах
имеются десятки последовательных синаптических контактов, становится понятной длительность большинства рефлекторных реакций — десятки миллисекунд.
Высокая утомляемость. Длительное повторное раздражение рецептивного поля рефлекса приводит к ослаблению рефлекторной реакции вплоть до полного исчезновения, что называется утомлением. Этот процесс связан с деятельностью синапсов — в последних наступает истощение запасов медиатора, уменьшаются энергетические ресурсы, происходит адаптация постсинаптического рецептора к медиатору.
6. Тонус. Тонус, или наличие определенной фоновой активности нервного центра, определяется тем, что в покое в отсутствие специальных внешних раздражений определенное количество нервных клеток находится в состоянии постоянного возбуждения, генерирует фоновые импульсные потоки. Даже во сне в высших отделах мозга остается некоторое количество фоновоактивных нервных клеток, формирующих «сторожевые пункты» и определяющих некоторый тонус соответствующего нервного центра.
7. Пластичность. Функциональная возможность нервного центра существенно модифицировать картину осуществляемых рефлекторных реакций. Поэтому пластичность нервных центров тесно связана с изменением эффективности или направленности связей между нейронами.
8. Конвергенция. Нервные центры высших отделов мозга являются мощными коллекторами, собирающими разнородную афферентную информацию. Количественное соотношение периферических рецепторных и промежуточных центральных нейронов (10:1) предполагает значительную конвергенцию («сходимость») разномодальных сенсорных посылок на одни и те же центральные нейроны. На это указывают прямые исследования центральных нейронов: в нервном центре имеется значительное количество поливалентных, полисенсорных нервных клеток, реагирующих на разномодальные стимулы (свет, звук, механические раздражения и т. д.). Конвергенция на клетках нервного центра разных афферентных входов предопределяет важные интегративные, перерабатывающие информацию функции центральных нейронов, т. е. высокий уровень интеграционных функций. Конвергенция нервных сигналов на уровне эфферентного звена рефлекторной дуги определяет физиологический механизм принципа «общего конечного пути» по Ч. Шеррингтону.
9. Интеграция в нервных центрах. Важные интегративные функции клеток нервных центров ассоциируются с интегративными процессами на системном уровне в плане образования функциональных объединений отдельных нервных центров в целях осуществления сложных координированных приспособительных целостных реакций организма (сложные адаптивные поведенческие акты).
10. Свойство доминанты. Доминантным называется временно господствующий в нервных центрах очаг (или доминантный центр) повышенной возбудимости в центральной нервной системе. По А.А.Ухтомскому, доминантный нервный очаг характеризуется такими свойствами, как повышенная возбудимость, стойкость и инертность возбуждения, способность к суммированию возбуждения.
В доминантном очаге устанавливается определенный уровень стационарного возбуждения, способствующий суммированию ранее подпороговых возбуждений и переводу на оптимальный для данныхусловий ритм работы, когда этот очаг становится наиболее чувствительным. Доминирующее значение такого очага (нервного центра) определяет его угнетающее влияние на другие соседние очаги возбуждения. Доминантный очаг возбуждения «притягивает» к себе возбуждение других возбужденных зон (нервных центров). Принцип доминанты определяет формирование главенствующего (активирующего) возбужденного нервного центра в тесном соответствии с ведущими мотивами, потребностями организма в конкретный момент времени.
11. Цефализация нервной системы. Основная тенденция в эволюционном развитии нервной системы проявляется в перемещении, сосредоточении функции регуляции и координации деятельности организма в головных отделах ЦНС. Этот процесс называется цефализацией управляющей функции нервной системы. При всей сложности складывающихся отношений между старыми, древними и эволюционно новыми нервными образованиями стволовой части мозга общая схема взаимных влияний может быть представлена следующим образом: восходящие влияния (от нижележащих «старых» нервных структур к вышележащим «новым» образованиям) преимущественно носят возбуждающий стимулирующий характер, нисходящие (от вышележащих «новых» нервных образований к нижележащим «старым» нервным структурам) носят угнетающий тормозной характер. Эта схема согласуется с представлением о росте в процессе эволюции роли и значения тормозных процессов в осуществлении сложных интегративных рефлекторных реакций.
42
Доминанта в физиологии, очаг возбуждения в центральной нервной системе, временно определяющий характер ответной реакции организма на внешние и внутренние раздражения. Доминирующий нервный центр (или группа центров) обладает повышенной возбудимостью и способностью стойко удерживать это состояние и тогда, когда первоначальный стимул уже не оказывает активирующего воздействия (инертность). Суммируя относительно слабые возбуждения др. центров, Д. одновременно воздействует на них тормозящим образом. В естественных условиях Д. образуется под влиянием рефлекторного возбуждения или при действии на нервные центры ряда гормонов. В эксперименте Д. может быть создана непосредственным воздействием на нервные центры слабым электрическим током или некоторыми фармакологическими веществами. Доминирование одних нервных центров над другими впервые описал Н. Е. Введенский (1881). Выясняя механизмы образования условных рефлексов, И. П. Павлов отмечал, что длительно поддерживаемый уровень повышенной возбудимости тех или иных участков коры больших полушарий определяет в значительной мере динамику высшей нервной деятельности в норме и патологии. Основные положения учения о Д. как общем принципе работы нервных центров сформулировал А. А. Ухтомский на основании выполненных им и его сотрудниками экспериментальных исследований (1911≈23). Д. выражается в готовности определённого органа к работе и поддержании его рабочего состояния. Д. в высших центрах головного мозга служит физиологической основой ряда психических явлений (например, внимания и др.). ═ Лит.: Ухтомский А. А., Доминанта, М.≈Л., 1966; Механизмы доминанты. (Материалы симпозиума), Л., 1967. ═ Н. Г. Алексеев, М. Ю. Ульянов.
43-44
Торможение в центральной нервной системе
Интегративная и координационная деятельность центральных нервных образований осуществляется при обязательном участии тормозных процессов.
Торможение в центральной нервной системе — активный процесс, проявляющийся внешне в подавлении или в ослаблении процесса возбуждения и характеризующийся определенной интенсивностью и длительностью.
Торможение в норме неразрывно связано с возбуждением, является его производным, сопутствует возбудительному процессу, ограничивая и препятствуя чрезмерному распространению последнего. При этом торможение часто ограничивает возбуждение и вместе с ним формирует сложную мозаику активированных и заторможенных зон в центральных нервных структурах. Формирующий эффект тормозного процесса развивается в пространстве и во времени. Торможение — врожденный процесс, постоянно совершенствующийся в течение индивидуальной жизни организма.
При значительной силе фактора, вызвавшего торможение, оно может распространяться на значительное пространство, вовлекая в тормозной процесс большие популяции нервных клеток.
История развития учения о тормозных процессах в центральной нервной системе начинается с открытия И. М. Сеченовым эффекта центрального торможения (химическое раздражение зрительных бугров тормозит простые спинномозговые безусловные реакции). Вначале предположение о существовании специфических тормозных нейронов, обладающих способностью оказывать тормозные влияния на другие нейроны, с которыми имеются синаптические контакты, диктовалось логической необходимостью для объяснения сложных форм координационной деятельности центральных нервных образований. Впоследствии это предположение нашло прямое экспериментальное подтверждение (Экклс, Реншоу), когда было показано существование специальных вставочных нейронов, имеющих синаптические контакты с двигательными нейронами. Активация этих вставочных нейронов закономерно приводила к торможению двигательных нейронов. В зависимости от нейронного механизма, способа вызывания тормозного процесса в ЦНС различают несколько видов торможения: постсинаптическое, пресинаптическое, пессимальное.
Постсиналтическое торможение — основной вид торможения, развивающийся в постсинаптической мембране аксосоматических и аксодендритических синапсов под влиянием активации тормозных нейронов, в концевых разветвлениях аксонных отростков которых освобождается и поступает в синаптическую щель тормозной медиатор. Тормозной эффект таких нейронов обусловливается специфической природой медиатора — химического переносчика сигнала с одной клетки на другую. Наиболее распространенным тормозным медиатором является гамма-аминомасляная кислота (ГАМК). Химическое действие ГАМК вызывает в постсинаптической мембране эффект гиперполяризации в виде тормозных постсинаптических потенциалов (ТПСП), пространственно-временная суммация которых повышает уровень мембранного потенциала (гиперполяризация), приводит к урежению или полному прекращению генерации распространяющихся ПД.
Возвратным торможением называется угнетение (подавление) активности нейрона, вызываемое возвратной коллатералью аксона нервной клетки. Так, мотонейрон переднего рога спинного мозга прежде чем покинуть спинной мозг дает боковую (возвратную) ветвь, которая возвращается назад и заканчивается на тормозных нейронах (клетки Реншоу). Аксон последней заканчивается на мотонейронах, оказывая на них тормозное действие.
Пресинаптическое торможение развертывается в аксоаксональных синапсах, блокируя распространение возбуждения по аксону. Пресинаптическое торможение часто выявляется в структурах мозгового ствола, в спинном мозге
Пессимальное торможение представляет собой вид торможения центральных нейронов. Оно наступает при высокой частоте раздражения. В первый момент возникает высокая частота ответного возбуждения. Через некоторое время стимулируемый центральный нейрон, работая в таком режиме, переходит в состояние торможения.
45
Пресинаптическое торможение, как свидетельствует название, локализуется в пресинаптических элементах и связано с угнетением проведения нервных импульсов в аксональных (пресинаптических) окончаниях. Гистологическим субстратом такого торможения является аксональные синапсы. К возбуждающему аксону подходит вставной тормозной аксон, который выделяет тормозной медиатор ГАМК. Этот медиатор действует на постсинаптическую мембрану, которая является мембраной возбуждающего аксона, и вызывает в ней деполяризацию. Возникшая деполяризация тормозит вход Са2 + из синаптической щели в заключение возбуждающего аксона и таким образом приводит к снижению выброса возбуждающего медиатора в синаптическую щель, торможение реакции. Пресинаптическое торможение достигает максимума через 15-20 мс и длится около 150 мс, то есть гораздо дольше, чем постсинаптическое торможение. Пресинаптическое торможение блокируется судорожными ядами - бикулином и пикротоксин, которые являются конкурентными антагонистами ГАМК. Постсинаптическое торможение (ГПСП) обусловлено выделением пресинаптическим окончанием аксона тормозного медиатора, который снижает или тормозит возбудимость мембран сомы и дендритов нервной клетки, с которой он контактирует. Оно связано с существованием тормозных нейронов, аксоны которых образуют на соме и дендритах клеток нервных окончаний, выделяя тормозные медиаторы - ГАМК и глицин. Под влиянием этих медиаторов возникает торможение возбуждающих нейронов. Примерами тормозных нейронов являются клетки Реншоу в спинном мозге, нейроны грушевидные (клетки Пуркинье мозжечка), звездчатые клетки коры большого, мозга и др.. Исследованием П. Г. Костюка (1977) доказано, что постсинаптического торможения связано с первичной гиперполяризацией мембраны сомы нейрона, в основе которой лежит повышение проницаемости постсинаптической мембраны для К +. Вследствие гиперполяризации уровень мембранного потенциала удаляется от критического (порогового) уровня. То есть происходит его увеличение - гиперполяризация. Это приводит к торможению нейрона. Такой вид торможения называется гиперполяризационным. Амплитуда и полярность ГПСП зависят от исходного уровня мембранного потенциала самого нейрона. Механизм этого явления связан с Сl+. С началом развития ТПСП Сl- входит в клетку. Когда в клетке становится его больше, чем снаружи, глицин конформирует мембрану и через открытые ее отверстия Сl+ выходит из клетки. В ней уменьшается количество отрицательных зарядов, развивается деполяризация. Такой вид торможения называется деполяризационным.
46
Принципы координации в деятельности центральной нервной системы
В условиях физиологической нормы работа всех органов и систем тела является согласованной: на воздействия из внешней и внутренней среды организм реагирует как единое целое. Согласованное проявление отдельных рефлексов, обеспечивающих выполнение целостных рабочих актов, носит название координации.
Явления координации играют важную роль в деятельности двигательного аппарата. Координация таких двигательных актов, как ходьба или бег, обеспечивается взаимосвязанной работой нервных центров.
За счет координированной работы нервных центров осуществляется совершенное приспособление организма к условиям существования. Это происходит не только за счет деятельности двигательного аппарата, но и за счет изменений вегетативных функций организма (процессов дыхания, кровообращения, пищеварения, обмена веществ и т. д.).
Установлен ряд общих закономерностей - принципов координации: 1) принцип конвергенции; 2) принцип иррадиации возбуждения; 3) принцип реципрокности; 4) принцип последовательной смены возбуждения торможением и торможения возбуждением; 5) феномен "отдачи"; 6) цепные и ритмические рефлексы; 7) принцип общего конечного пути; 8) принцип обратной связи; 9) принцип доминанты.
Разберем некоторые из них.
Принцип конвергенции. Этот принцип установлен английским физиологом Шеррингтоном. Импульсы, приходящие в центральную нервную систему по различным афферентным волокнам, могут сходиться (конвергировать) к одним и тем же вставочным и эфферентным нейронам. Конвергенция нервных импульсов объясняется тем, что афферентных нейронов в несколько раз больше, чем эфферентных, поэтому афферентные нейроны образуют на телах и дендритах эфферентных и вставочных нейронов многочисленные синапсы.
Принцип иррадиации. Импульсы, поступающие в центральную нервную систему при сильном и длительном раздражении рецепторов, вызывают возбуждение не только данного рефлекторного центра, но и других нервных центров. Это распространение возбуждения в центральной нервной системе получило название иррадиации. Процесс иррадиации связан с наличием в центральной нервной системе многочисленных ветвлений аксонов и особенно дендритов нервных клеток и цепей вставочных нейронов, которые объединяют друг с другом различные нервные центры.
Принцип реципрокности (сопряженности) в работе нервных центров. Это явление было изучено И. М. Сеченовым, Н. Е. Введенским, Шеррингтоном. Суть его заключается в том, что при возбуждении одних нервных центров деятельность других может затормаживаться. Принцип реципрокности был показан по отношению к нервным центрам мышц-антагонистов - сгибателей и разгибателей конечностей. Наиболее отчетливо он проявляется у животных с удаленным головным мозгом и сохраненным спинным (спинальное животное). Если раздражать у спинального животного (кошка) кожу конечностей, отмечается сгибательный рефлекс данной конечности, а на противоположной стороне в это время наблюдается рефлекс разгибания. Описанные явления связаны с тем, что при возбуждении центра сгибания одной конечности происходит реципрокное торможение центра разгибания этой же конечности. На симметричной стороне имеются обратные взаимоотношения: возбужден центр разгибателей и заторможен центр сгибателей. Только при такой взаимосочетанной (реципрокной) иннервации возможен акт ходьбы.
Может происходить сопряженное, реципрокное торможение и других рефлексов. Под влиянием головного мозга реципрокные отношения могут изменяться. Человек или животное в случае необходимости может сгибать обе конечности, совершать прыжки и т. д.
Реципрокные взаимоотношения центров головного мозга определяют возможность человека овладеть сложными трудовыми процессами и не менее сложными специальными движениями, совершающимися при плавании, акробатических упражнениях и пр.
Принцип общего конечного пути. Этот принцип связан с особенностью строения центральной нервной системы. Эта особенность, как уже указывалось, состоит в том, что афферентных нейронов в несколько раз больше, чем эфферентных, в результате чего различные афферентные импульсы сходятся к общим выходящим путям. Количественные соотношения между нейронами схематически можно представить в виде воронки: возбуждение вливается в центральную нервную систему через широкий раструб (афферентные нейроны) и вытекает из нее через узкую трубку (эфферентные нейроны). Общими путями могут быть не только конечные эфферентные нейроны, но и вставочные.
Импульсы, сходящиеся в общем пути, "конкурируют" друг с другом за использование этого пути. Так достигается упорядочение рефлекторного ответа, соподчинение рефлексов и затормаживание менее существенных. Вместе с тем организм получает возможность реагировать на различные раздражения из внешней и внутренней среды при помощи сравнительно небольшого количества исполнительных органов.
Принцип обратной связи. Этот принцип изучен И. М. Сеченовым, Шеррингтоном, П. К. Анохиным и рядом других исследователей. При рефлекторном сокращении скелетных мышц возбуждаются проприорецепторы. От проприорецепторов нервные импульсы вновь поступают в центральную нервную систему. Этим контролируется точность совершаемых движений. Подобные афферентные импульсы, возникающие в организме в результате рефлекторной деятельности органов и тканей (эффекторов), получили название вторичных афферентных импульсов, или обратной связи.
Обратные связи могут быть положительными и отрицательными. Положительные обратные связи способствуют усилению рефлекторных реакций, отрицательные - их угнетению.
За счет положительных и отрицательных обратных связей осуществляется, например, регуляция относительного постоянства величины артериального давления.
При повышении артериального давления происходит возбуждение механорецепторов дуги аорты, каротидных синусов. Импульсы поступают в сосудодвигательный центр и центр сердечной деятельности, тонус сосудов рефлекторно снижается, одновременно замедляется деятельность сердца и величина артериального давления уменьшается. При понижении артериального давления раздражение механорецепторов указанных рефлексогенных зон вызывает рефлекторное повышение тонуса сосудов, увеличение работы сердца. В этом случае величина артериального давления возрастает.
Вторичные афферентные импульсы (обратные связи) играют также важную роль в регуляции других вегетативных функций: дыхания, пищеварения, выделения.
Принцип доминанты. Принцип доминанты сформулирован А. А. Ухтомским. Этот принцип играет важную роль в согласованной работе нервных центров. Доминанта - это временно господствующий очаг возбуждения в центральной нервной системе, определяющий характер ответной реакции организма на внешние и внутренние раздражения.
Доминантный очаг возбуждения характеризуется следующими основными свойствами: 1) повышенной возбудимостью; 2) стойкостью возбуждения; 3) способностью к суммированию возбуждения; 4) инерцией - доминанта в виде следов возбуждения может длительно сохраняться и после прекращения вызвавшего ее раздражения.
Доминантный очаг возбуждения способен притягивать (привлекать) к себе нервные импульсы из других нервных центров, менее возбужденных в данный момент. За счет этих импульсов активность доминанты еще больше увеличивается, а деятельность других нервных центров подавляется.
Доминанты могут быть экзогенного и эндогенного происхождения. Экзогенная доминанта возникает под влиянием факторов окружающей среды. Например, при чтении интересной книги человек может не слышать звучащую в это время по радио музыку.
Эндогенная доминанта возникает под влиянием факторов внутренней среды организма, главным образом гормонов и других физиологически активных веществ. Например, при понижении содержания питательных веществ в крови, особенно глюкозы, происходит возбуждение пищевого центра, что является одной из причин пищевой установки организма животных и человека.
Доминанта может быть инертной (стойкой), и для ее разрушения необходимо возникновение нового более мощного очага возбуждения.
Доминанта лежит в основе координационной деятельности организма, обеспечивая поведение человека и животных в окружающей среде, а также эмоциональных состояний, реакций внимания. Формирование условных рефлексов и их торможение также связано с наличием доминантного очага возбуждения.
47
Вегетативная нервная система.
Вегетативная (автономная) нервная система — регулирует деятельность внутренних органов, обеспечивает важнейшие функции питания, дыхания, выделения, размножения, циркуляции крови и лимфы. Ее реакции не подчинены напрямую нашему сознанию компоненты вегетативной нервной системы пронизывают практически все ткани организма, вместе с гормонами желез внутренней секреции (эндокринных желез) она координирует работу органов, подчиняя ее общей цели — созданию оптимальных условий существования организма в данной ситуации и в данный момент времени.
Нервные клетки вегетативной нервной системы находятся не только в головном и спинном мозге, они широко рассеяны во многих органах, особенно в желудочно-кишечном тракте. Они в виде многочисленных узлов (ганглиев) располагаются между органами и мозгом. Вегетативные нейроны образуют друг с другом связи, позволяющие им работать автономно, образуется масса мелких нервных центров вне пределов центральной нервной системы, которые могут взять на себя некоторые относительно простые функции (например, организацию волнообразных сокращений кишечника). При этом центральная нервная система продолжает осуществлять общий контроль за ходом этих процессов и вмешиваться в них.
В вегетативной нервной системе выделяют симпатическую и парасимпатическую части. При преобладающем влиянии одной из них орган снижает или, наоборот, усиливает свою работу. Обе они находятся под контролем высших отделов центральной нервной системы, чем достигается их согласованное действие. Вегетативные центры в головном и спинном мозге составляют центральный отдел вегетативной нервной системы, а ее периферический отдел представлен нервами, узлами, вегетативными нервными сплетениями.
Симпатические центры расположены в боковых рогах серого вещества спинного мозга, в его грудных и поясничных сегментах. От их клеток отходят симпатические волокна, которые в составе передних корешков, спинномозговых нервов и их веточек направляются к узлам симпатического ствола. Правый и левый симпатические стволы расположены вдоль всего позвоночного столба. Они представляют собой цепочку утолщений (узлов), в которых находятся тела симпатических нервных клеток. К ним и подходят нервные волокна от центров спинного мозга. Отростки же клеток узлов идут к внутренним органам в составе вегетативных нервов и сплетений.
Симпатические стволы имеют шейный, грудной, поясничный и тазовый отделы. Шейный отдел состоит из трех узлов, ветви которых образуют сплетения на сосудах головы, шеи, груди, около органов и в их стенках, в том числе, сердечные сплетения. Грудной отдел включает 10-12 узлов, их веточки образуют сплетения на аорте, бронхах, в пищеводе. Проходя через диафрагму, они входят в состав солнечного сплетения. Поясничный отдел симпатического ствола образуют 3-5 узлов. Их ветви через солнечное и другие вегетативные сплетения брюшной полости достигают желудка, печени, кишечника, почек, поджелудочной железы, половых желез. Тазовый отдел включает 4 узла, через которые и осуществляется симпатический контроль над органами малого таза (мочевым пузырем, прямой кишкой).
Парасимпатические центры расположены в стволе головного мозга и в крестцовых сегментах спинного мозга. Отростки их нейронов идут, как правила, непосредственно до органов, а уже в их стенках находятся тела последних в этой цепочке нервных клеток с очень короткими отростками. Парасимпатические центры мозгового ствола через черепные нервы контролируют органы головы, шеи, а посредством блуждающего нерва — и органов грудной и брюшной полостей. Волокна от крестцовых центров идут по тазовым вегетативным сплетениям к органам таза и брюшной полости. Антагонизм и синергизм — две стороны единого процесса. Нормальные функции нашего организма обеспечиваются согласованным действием этих двух отделов вегетативной нервной системы. Эта согласованность и регуляция функций осуществляются корой головного мозга. В этой регуляции участвует и ретикулярная формация.
Вегетативная нервная система отличается от соматической следующими признаками: 1. Вегетативный отдел нервной системы иннервирует гладкую мускулатуру, которая сокращается непроизвольно (без контроля сознания), медленно, а в отдельных органах находится в состоянии длительного сокращения (сфинктеры полых органов) или постоянного сокращения (гладкие мышцы стенок кровеносных сосудов). Соматический отдел иннервирует скелетную мускулатуру и побуждает её к выполнению быстрых и целенаправленных сокращений, осуществляющихся под контролем сознания. 2. Кроме того, что вегетативный отдел нервной системы иннервирует гладкую мускулатуру внутренних органов и сосудов, мышцу сердца и железы, она обеспечивает трофическую иннервацию всех тканей и органов, включая скелетную мускулатуру, т.е. отличается от соматического отдела повсеместностью распространения. 3. Важнейший отличительный признак вегетативного отдела - это очаговый характер расположения центров (ядер) в стволе головного мозга (мезенцефалический и бульбарный отделы) и спинном мозге (тораколюмбальный и сакральный отделы). Соматические же центры располагаются в пределах центральной нервной системы равномерно (сегментарно). 4. Очаговость расположения центров предопределяет и очаговый характер выхода корешков вегетативных волокон от мозгового ствола и спинного мозга. Они выходят в составе III, YII, IX, X пар черепных и C8 - L3, S2 - S4 спинномозговых нервов. При этом принцип строгой сегментарности отсутствует не только в месте отхождения, но и в распределении волокон на периферии. 5. Различия в строении рефлекторной дуги (см. выше). 6. Деятельность вегетативной нервной системы основана не только на центральных рефлекторных дугах, но и на периферических, двухнейронных, замыкающихся в вегетативных узлах. 7. Вегетативная нервная система сохранила ряд примитивных черт в своём строении: а) диффузность локализации нейронов в виде ганглиев; б) сравнительное однообразие форм и размеров нейронов, нервные клетки как правило мелкие, многоотростчатые (рис. 3); в) меньший калибр нервных волокон из-за тонкой миелиновой оболочки (преганглионарные волокна) или ее отсутствия (постганглионарные волокна), что обусловливает низкую возбудимость и скорость проведения импульса. В соматических афферентных волокнах скорость составляет 70-120 м/сек, в вегетативных - 0,3-10 м/сек. 8. Вегетативная нервная система обладает избирательной чувствительностью к гормонам и различного рода метаболитам. Это обусловлено тем, что переключение импульса в синапсах осуществляется с помощью химического вещества - медиатора. Некоторые биологически активные вещества и гормоны являются стимуляторами в механизмах освобождения и выделения медиаторов и тем самым повышают тонус симпатического или парасимпатического отдела вегетативной нервной системы.
48
Регуляция функций вегетативной нервной системой осуществляется по рефлекторному принципу. Т.е. раздражение периферических рецепторов приводит к возникновению нервных импульсов, которые после анализа и синтеза в вегетативных центрах поступают на эфферентные нейроны, а затем исполнительные органы. Поэтому все вегетативные рефлексы, в зависимости от участия рецепторного и эффекторного, звена делятся на следующие группы:
1.Висцеро-висцеральные. Это рефлексы, которые возникают вследствие раздражения интерорецепторов внутренних органов и проявляются изменениями их функций. Например, при механическом раздражении брюшины или органов брюшной полости происходит урежение и ослабление сердечных сокращений. Рефлекс Гольца.
2.Висцеро-дермальные. Раздражении интерорецепторов внутренних органов, приводит к изменению потоотделения, просвета сосудов кожи, кожной чувствительности.
3.Сомато-висцеральные. Действие раздражителя на соматические рецепторы, например рецепторы кожи, приводит к изменению деятельности внутренних органов. К этой группе относится рефлекс Данини-Ашнера.
4.Висцеро-соматические. Раздражение интерорецепторов вызывает изменение двигательных функций. Возбуждение хеморецепторов сосудов углекислым газом, способствует усилению сокращений межреберных дыхательных мышц.
При нарушении механизмов вегетативной регуляции возникают изменения висцеральных функций. В частности, психосоматические заболевания.
49
Симпатическая нервная система – часть вегетативной нервной системы, регулирующая совместно с парасимпатической нервной системой деятельность внутренних органов и обмен веществ в организме. Анатомические образования, составляющие симпатическую нервную систему, располагаются как в центральной нервной системе, так и вне её. Спинальные симпатические центры находятся под контролем высших вегетативных нервных центров, расположенных в головном мозге. От этих симпатических центров идут симпатические нервные волокна, которые, выйдя из спинного мозга с передними мозговыми корешками, попадают в пограничный симпатический ствол (левый и правый), расположенный параллельно позвоночнику.
Каждый узел симпатического ствола связан с определёнными отделами организма и внутренними органами через нервные сплетения. Из грудных узлов выходят волокна, образующие солнечное сплетение, из нижних грудных и верхних поясничных - почечное сплетение. Практически каждый орган имеет собственное сплетение, образующееся путём дальнейшего разделения указанных крупных симпатических сплетений и их соединения с подходящими к органам парасимпатическим волокнами. От сплетений, где происходит передача возбуждения с одной нервной клетки на другую, симпатические волокна подходят непосредственно к органам, мышцам, сосудам и тканям. Передача возбуждения с симпатического нерва на рабочий орган осуществляется с помощью определённых химических веществ (медиаторов) – симпатинов, выделяющихся нервными окончаниями. По своему химическому составу симпатины близки к гормону мозгового слоя надпочечников – адреналину.
При раздражении симпатических нервных волокон большинство периферических кровеносных сосудов (за исключением сосудов сердца, обеспечивающих нормальное питание сердца) суживается, ритм сердечных сокращений учащается, расширяются зрачки, выделяется густая вязкая слюна и так далее. Отмечается выраженное влияние симпатической нервной системы на ряд процессов обмена веществ, одним из проявлений которого является повышение уровня сахара в крови, повышенное теплообразование и уменьшение теплоотдачи, увеличение свёртываемости крови.
Нарушения деятельности симпатической нервной системы могут возникнуть в результате инфекционного или токсического поражения её образований. При нарушении функции симпатической нервной системы могут наблюдаться местные и общие расстройства кровообращения, расстройства деятельности пищеварительного аппарата, нарушение сердечной деятельности, нарушения питания тканей. Повышенная возбудимость симпатической нервной системы обнаруживается при таких распространённых заболеваниях, как, например, гипертоническая и язвенная болезни, неврастения и другие.
Лечение нарушений симпатической нервной системы требует прежде всего борьбы с факторами, лежащими в их основе, а так же применения лекарственных веществ, воздействующих на различные отделы вегетативной нервной системы.
50
Парасимпатическая нервная система, часть вегетативной нервной системы, ганглии которой расположены в непосредственной близости от иннервируемых органов или в них самих. Центры П. н. с. находятся в среднем и продолговатом мозге (мезенцефальный и бульбарный отделы), а также в крестцовом участке спинного мозга (сакральный отдел). Волокна П. н. с. направляются к внутренним органам в составе глазодвигательного (III пара), лицевого (VII пара), языкоглоточного (IX пара) и главным образом блуждающего (Х пара) черепномозговых нервов, а также в составе тазового нерва. Во многих случаях воздействия на органы со стороны П. н. с. и симпатической нервной системы (СНС) прямо противоположны друг другу. Так, если под влиянием импульсов, поступающих по симпатическим нервам, учащаются и усиливаются сокращения сердца, повышается кровяное давление, расширяется зрачок, то импульсы, приходящие по волокнам П. н. с., вызывают замедление и ослабление сердцебиений, понижают артериальное давление, суживают зрачок. Последний, эфферентный, нейрон П. н. с. расположен, как правило, в самом иннервируемом органе, а не в т. н. пограничном стволе, как это имеет место в СНС. Медиатором, образующимся в нервных окончаниях П. н. с., является ацетилхолин (преобладающий медиатор в СНС — норадреналин).
51
11.1. Метасимпатическая нервная система
Метасимпатическая (А.Д. Ноздрачёв), или энтеральная (Ленгли) нервная система является периферической, эволюционно самой древней частью вегетативной системы. У человека количество входящих в неё нейронов значительно превышает количество нейронов симпатической и парасимпатической систем; их около 108, то есть примерно столько же, сколько в спинном мозге.
Нейроны метасимпатической системы рассеяны по всему организму. Во внутренних органах (желудочно-кишечный тракт, дыхательная, сердечно-сосудистая и мочеполовая система) они образуют интрамуральные (встроенные) нервные сети (сплетения), регулирующие работу этих органов за счёт местных рефлексов. Наибольшее число нейронов метасимпатической системы располагается в межмышечном (ауэрбаховом) и подслизистом (мейснеровском) сплетениях желудочно-кишечного тракта.
Вегетативная рефлекторная дуга метасимпатической системы состоит из сенсорных, вставочных и двигательных нейронов, которые за счёт многочисленных отростков контактируют с большим числом других клеток, образующих, в свою очередь, разные звенья других дуг.
Такая диффузная сеть нейронов реагирует как единое целое на местные стимулы. Например, воспринимаемое афферентными нейронами растяжение гладкой мускулатуры стенки кишечника или химическое раздражение слизистой оболочки, изменяет активность выше и нижележащих вставочных и эфферентных нейронов и согласовывает сократительную и секреторную активность участков кишечника, расположенных выше и ниже места стимуляции, а также непосредственно стимулируемого участка. В результате согласованного сокращения мускулатуры кишечника осуществляется продвижение пищевых масс со скоростью, оптимальной для их расщепления и Виды нейронов по медиаторам синапса
серотонинергические
холинергические
пуринергические
андренергические
пептидергические
возможно, гистаменергические
52
Вегетативная нервная система имеет центральный и периферический отделы. В центральном отделе различают надсегментарные (высшие) и сегментарные (низшие) вегетативные центры. Надсегментарные вегетативные центры сосредоточены в головном мозге — в коре головного мозга (преимущественно в лобных и теменных долях), гипоталамусе, обонятельном мозге, подкорковых структурах (полосатое тело), в стволе головного мозга (ретикулярная формация), мозжечке и др. Сегментарные вегетативные центры расположены и в головном, и в спинном мозге. Вегетативные центры головного мозга условно подразделяют на среднемозговые и бульбарные (вегетативные ядра глазодвигательного, лицевого, языко-глоточного и блуждающего нервов), а спинного мозга — на пояснично-грудинные и крестцовые (ядра боковых рогов сегментов CVIII—LIII и SII—SIV соответственно).
Моторные центры иннервации неисчерченных (гладких) мышц внутренних органов и сосудов расположены в предцентральной и лобной областях. Здесь же находятся центры рецепции из внутренних органов и сосудов, центры потоотделения, нервной трофики, обмена веществ. В полосатом теле сосредоточены центры терморегуляции, слюно- и слезоотделения. Установлено участие мозжечка в регуляции таких вегетативных функций, как зрачковый рефлекс, трофика кожи. Ядра ретикулярной формации составляют надсегментарные центры жизненно важных функци�� — дыхательной, сосудодвигательной, сердечной деятельности, глотания и др.
Периферический отдел В. н. с. представлен нервами и узлами, расположенными вблизи внутренних органов (экстрамурально) либо в их толще (интрамурально). Вегетативные узлы соединяются между собой нервами, образуя сплетения, например легочное, сердечное, брюшное аортальное сплетение.
(в книге)
53
3.9. ТРОФИЧЕСКОЕ ДЕЙСТВИЕ НЕРВНОЙ СИСТЕМЫ
Идею о трофическом действии НС сформулировал И.П. Павлов. В результате опыта на собаке он обнаружил симпатическую веточку, идущую к сердцу, раздражение которой вызывает усиление сердечных сокращений без изменения их частоты (усиливающий нерв Павлова). Впоследствии было показано, что раздражение симпатического нерва, действительно, усиливает в сердце обменные процессы. Развивая идею И. П. Павлова, Л. А. Орбели и А. Г. Гинецинский в 20-х годах (XX в.) обнаружили усиление сокращений утомленной скелетной мышцы при раздражении идущего к ней симпатического нерва (феномен Орбели - Гинецинского, рис. 3.5). Считается, что усиление сокращений утомленной мышцы в опыте Орбели - Г инецинского связано с активацией в ней обменных (трофических) процессов под влиянием норадреналина.
В дальнейшем было установлено, что раздражение симпатических нервов не только улучшает функциональные характеристики скелетных мышц, но повышает возбудимость периферических рецепторов и структур ЦНС. Такое действие симпатической нервной системы Л.А. Орбели назвал адаптационно-трофическим. Трофическое действие на ткань присуще всем нервам, но наиболее ярко оно выражено у симпатической НС. Предполагается наличие трофогенов в нервных окончаниях. К трофогенам относят нукле-отиды, некоторые аминокислоты, простагландины, катехоламины, серотонин, ацетилхолин, сложные липиды, ганглиозиды. Многие из перечисленных веществ являются медиаторами - понятие «тро-фоген», по-видимому, является собирательным.
Трофическое действие на иннервируемые ткани оказывают и афферентные нервные волокна, что осуществляется преимущественно нейропептидами, например субстанцией Р.