

5)Хранение в dist. Или 1%-ном NaCl.Мо предварительно выращивают в оптимальных условиях, после чего клетки суспендируют в дистиллированной воде или 1%-ном растворе хлорида натрия.
6)Хранение в высушенном состоянии на адсорбентах.применяют главным образом для актиномицетов,анаэробных спорообраз.бактерий. В качестве адсорбентов используют почву, кварцевый песок, силикагель, вату, фильтровальную бумагу.Стерильный адсорбент, помещенный в ампулы, смешивают с густой суспензией клеток и высушивают под вакуумом или при комнатной температуре. Затем ампулы запаивают и хранят при комнатной температуре или в холодильнике.
Билет №24
Вопрос 1. Круговорот серы
В живых клетках S представлена главным образом сульфгидрильными группами в серусодержащих а/к (Cys, Met, гомоCys). В сухом веществе организмов доля серы составляет 1%. При анаэробном разложении органических веществ сульфгидрильные группы отщепляются десульфуразами; образование H2S при минерализации в анаэробных условиях называют также десульфурированием. Наибольшие количества встречающегося в природе H2S образуются, однако, при диссимиляционном восстановлении сульфатов, осуществляемом сульфатредуцирующими бактериями. Этот сероводород, образующийся в отсутствие молекулярного кислорода в осадках водоемов, может быть окислен анаэробными фототрофными бактериями до серы и сульфата. Когда сероводород проникает в зоны, содержащие 02, он окисляется либо абиотическим образом, либо аэробными серобактериями до сульфата. Серу, необходимую для синтеза серусодержащих а/к, растения и часть мо получают путем ассимиляционной сульфатредукции; животные же получают восстановленные соединения серы с пищей.
Вопрос 2. Принцип работы лактозного оперона
Вопрос-принцип работы лактозного оперона
Р егуляторный ген |
|
САР |
помотор |
о перон |
Структурный ген 1βгалактоза |
Структурный ген2пермиаза |
Структурный ген 3тиогалактоза |
Регуляторный ген синтезирует белок, который взаимодействует с оператором.
Ген1 ответственен за расщепление галактозы,
Ген2ответственен за перенос , проникновение галактозы
Ген3 превращает галактозу в глюкозу.
Позитивный контроль:
глюкозы нет, есть комплекс цАМФ+САРбелок,
Глюкоза есть , нет цАМФ+САРбелок,
Негативный контроль:
Если нет глюкозы в организме, то комплекс цАМФ+САР обеспечивает посадку полимеразы
Если появляется лактоза, то регуляторный белок уходит с оператора и синтез становится возможным.
Лактозный оперон Е. coli, состоящий из трех структурных генов, промотора и оператора, был первой ферментной системой, на которой Ж. Моно и Ф. Жакоб изучали механизм индукции синтеза ферментов.
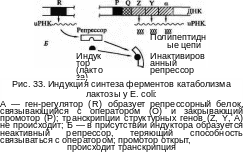 В отсутствие
лактозы молекула репрессора, активная
в свободном состоянии, связывается с
оператором и подавляет транскрипцию
структурных генов. Когда в клетку
попадает лактоза, она связывается с
репрессором, в результате образуется
неактивный комплекс репрессора с
индуктором, который не может
взаимодействовать с оператором и,
следовательно, препятствовать транскрипции
структурных генов. В результате
индуцируется синтез ферментов катаболизма
лактозы. При удалении из клетки индуктора
peпpeccop снова переходит в активное
свободное состояние, связывается с
оператором, что приводит к прекращению
синтеза соответствующих ферментов.
В отсутствие
лактозы молекула репрессора, активная
в свободном состоянии, связывается с
оператором и подавляет транскрипцию
структурных генов. Когда в клетку
попадает лактоза, она связывается с
репрессором, в результате образуется
неактивный комплекс репрессора с
индуктором, который не может
взаимодействовать с оператором и,
следовательно, препятствовать транскрипции
структурных генов. В результате
индуцируется синтез ферментов катаболизма
лактозы. При удалении из клетки индуктора
peпpeccop снова переходит в активное
свободное состояние, связывается с
оператором, что приводит к прекращению
синтеза соответствующих ферментов.
Лактозный оперон (lac operon) состоит из трех структурных генов, промотора, оператора и терминатора. Принимается, что в состав оперона входит также ген-регулятор, который кодирует белок-репрессор.
Структурные гены лактозного оперона — lacZ, lacY и lacA:
lacZ кодирует фермент β-галактозидазу, которая расщепляет дисахарид лактозу на глюкозу и галактозу,
lacY кодирует β-галактозид пермеазу, мембранный транспортный белок, который переносит лактозу внутрь клетки.
lacA кодирует β-галактозид трансацетилазу, фермент, переносящий ацетилную группу от ацетил-КoA на бета-галактозиды.
Для катаболизма лактозы необходимы только продукты генов lacZ и lacY, роль продукта гена lacA не ясна.
Регуляция(подробнее)
РНК-полимераза начинает транскрипцию с промоторного района, который перекрывается с операторным районом. В отсутствие или при низкой концентрации лактозы в клетке белок-репрессор, который является продуктом моноцистронного оперона LacI, обратимо соединяется с операторным районом и препятствует транскрипции. Таким образом, в отсутствие лактозы в клетке ферменты для метаболизма лактозы не синтезируются.
Даже в случае, когда в плазматической мембране клетки отсутствует фермент β-галактозидпермеаза, лактоза из окружающей среды может попадать в клетку в небольших количествах. В клетке две молекулы лактозы связываются с белком-репрессором, что приводит к изменению его конформации и далее к диссоциации белка-репрессора от операторного участка. Может осуществляться транскрипция генов лактозного оперона. При снижении концентрации лактозы новые порции белка-репрессора взаимодействуют с операторными последовательностями и препятствуют транскрипции. Данный механизм регуляции активности лактозного оперона называют позитивной индукцией. Веществом-индуктором служит лактоза; при её связывании с белком-репрессором происходит его диссоциация от операторного участка.
Если в клетке концентрация глюкозы достаточная для поддержания метаболизма, активация лактозного оперона не происходит. Промоторная последовательность лактозного оперона «слабая», поэтому даже при отсутствии белка-репрессора на операторном участке транскрипция практически не инициируется. Когда концентрация глюкозы в клетке снижается, происходит активация фермента аденилатциклазы, которая катализирует превращение АТР в циклическую форму — сАМР (циклическую форму АМР в данном случае также называют «сигналом клеточного голода»). Глюкоза является ингибитором фермента аденилатциклазы и активирует фосфодиэстеразу — фермент, катализирующий превращение молекулы сАМР в АМР. сАМР соединяется с белком, активирующим катаболизм (англ. САР, catabolism activating protein), при этом образуется комплекс, который взаимодействует с промотором лактозного оперона, изменяет его конформацию и приводит к повышению сродства РНК-полимеразы к данному участку. В присутствии лактозы происходит экспрессия генов оперона. Данный механизм регуляции активности лактозного оперона называют негативной индукцией. «Негативным индуктором» служит глюкоза, которая подавляет активность лактозного оперона.
Итак, ферменты для усвоения лактозы синтезируются в клетке кишечной палочки при двух условиях: 1) наличие лактозы; 2) отсутствие глюкозы. Регуляция работы лактозного оперона в зависимости от концентрации лактозы происходит по принципу отрицательной обратной связи: чем больше лактозы - тем больше ферментов для её катаболизма (положительная прямая связь); чем больше ферментов - тем меньше лактозы, чем меньше лактозы - тем меньше производится ферментов (двойная отрицательная обратная связь).