

Ферментативный катализ
Ферментативный катализ -биокатализ, ускорение химических реакций под влиянием ферментов . В основе жизнедеятельности лежат многочисленные химические реакции расщепления питательных веществ, синтеза необходимых организму химических соединений и трансформации их энергии в энергию физиологических процессов (работа мышц, почек, нервная деятельность и т.п.). Все эти реакции не могли бы происходить с необходимой для живых организмов скоростью, если бы в ходе эволюции не возникли механизмы их ускорения с помощью Ф. к. Эффективность Ф. к. достигается в результате того, что химическая реакция разбивается на ряд энергетически более лёгких промежуточных реакций, в которых участвует фермент. Важнейшая для Ф. к. реакция – образование первичного фермент-субстратного комплекса даёт выигрыш энергии, достаточный для ускорения процесса в целом.
Электронно-конформационные взаимодействия
Под конформационной динамикой (или подвижностью) понимаются относительные смещения белковых групп с амплитудами, заметно превышающими амплитуды валентных колебаний атомов. Изменение характеристик валентных колебаний с характерными временами порядка 0,1 пс и амплитудами до 0,1 А происходит непосредственно в акте изменения электронного состояния групп, что обсуждалось выше в связи с электронно-колебательным взаимодействием. В белках же имеется еще один уровень организации атомных движений - конформационные движения с амплитудами порядка 1 А. Эти движения характеризуются временами (релаксации) от сотен до миллионов пикосекунд. Времена релаксации конформационных движений в отличие от случая валентных колебаний резко увеличиваются при понижении температуры.
Роль конформационных движений также пока неизвестна во всех деталях. С одной стороны, спонтанные тепловые движения белковых групп по конформационным степеням свободы способствуют выходу системы в реакционноспособную конфигурацию. Так, например, для реакций туннельного переноса электрона важно устроить хорошее перекрывание электронных орбиталей между участниками процесса С другой стороны, после акта переноса электрона появляются новые заряженные центры, и изменяется баланс электростатических сил в молекуле белка. Это приводит и к изменению равновесных положений молекулярных групп, то есть к определенному изменению пространственной структуры белка или конформационному переходу
11. Биофизика механохимических процессов.
Основной надмолекулярной двигательной структурой мышечных волокон является саркомер. Саркомеры расположены в мышечном волокне последовательно один за другим.
Основной
надмолекулярной двигательной структурой
мышечных волокон является саркомер.
Саркомеры расположены в мышечном волокне
последовательно один за другим.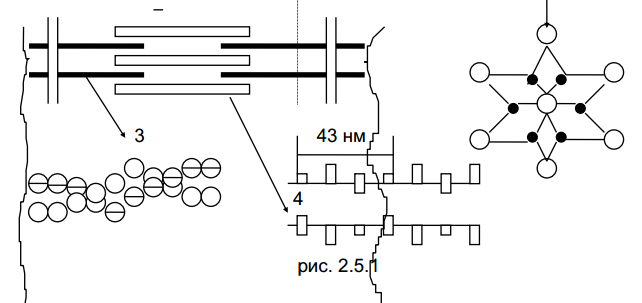
Величина
механического напряжения , возникающего
при сокращении мышцы (в определенных
пределах прямо пропорциональна
относительной деформации, согласно
закону Гука. Е = F/S - механическое
напряжение, F - развиваемая сила, S -
площадь поперечного сечения;
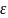 =
L/L0 - относительная деформация, L0
-первоначальная длина, L - абсолютная
деформация (удлинение); Е - модуль
упругости (модуль Юнга). Он определяется
величиной механического напряжения в
сокращенной мышце, при увеличении ее
длинны в два раза.
=
L/L0 - относительная деформация, L0
-первоначальная длина, L - абсолютная
деформация (удлинение); Е - модуль
упругости (модуль Юнга). Он определяется
величиной механического напряжения в
сокращенной мышце, при увеличении ее
длинны в два раза.

Функциональная роль АТФ в процессе сокращения скелетной мышцы.
- В результате вызываемого миозином гидролиза АТФ поперечные мостики получают энергию для развития тянущего усилия.
- Связывание АТФ с миозином сопровождается отсоединением поперечных мостиков , прикрепленных к актину , и создается возможность повторения цикла их активности.
- Гидролиз АТФ под действием Са2+-АТФазы саркоплазматического ретикулума поставляет энергию для активного транспорта Са2+ в латеральные цистерны саркоплазматического ретикулума, что приводит к снижению цитоплазматического Са2+ до исходного уровня; соответственно, сокращение завершается и мышечное волокно расслабляется.
