

Д. Клеточный цикл и его регуляция
Процессы роста и деления клеток лежат в основе жизни любого организма. Но прежде чем совершить деление, клетка должна с высокой точностью копировать свой геном, синтезировать множество высоко- и низкомолекулярных соединений. Совокупность событий, обеспечивающих деление эукариотических клеток, называют "клеточный цикл". Продолжительность клеточного цикла зависит от типа делящихся клеток, у взрослого человека она может варьировать примерно от 8 ч и более, а для некоторых типов клеток до года и больше (рис. 4-20).
Все фазы клеточного цикла G1, S, G2, M могут различаться по длительности, но в особенности это касается фазы G1, длительность которой может быть равна практически нулю или быть столь продолжительной, что может казаться, будто клетки вообще прекратили деление. В этом случае говорят, что клетки находятся в состоянии покоя (фаза G0). Так, нейроны взрослого человека не делятся вообще. Клетки эпителия кишечника делятся на протяжении всей жизни человека, но даже у этих быстропролиферирующих клеток подготовка к делению занимает 24 ч. Клетки лёгких, почек, печени во взрослом организме начинают делиться только лишь в ответ на повреждение органов.
Внешние сигналы могут стимулировать или ингибировать прохождение клетки через цикл. Пролиферативные сигналы очень разнообразны, они зависят от типа клетки, стадии развития и других факторов. Такими сигналами могут быть факторы роста, интерлейкины, гормоны, способные поддерживать или индуцировать пролиферацию определённых типов клеток. Сигнальные молекулы связываются специфическими мембранными рецепторами, активируют внутриклеточные пути передачи сигналов от рецептора к ядру и таким образом индуцируют транскрипцию определённых генов. Одними из первых активируются гены, кодирующие белки циклины. Белки были названы циклинами, потому что их концентрация в клетке периодически меняется по мере прохождения клеткой разных фаз клеточного цикла.
Все циклины делят на 2 подсемейства: G1-циклины (D, Е) и митотические циклины (А и В). Любой из циклинов представлен группой полиморфных белков, например циклин D представлен формами Dl, D2, D3. У каждого типа циклинов есть гомологичный участок из 100 аминокислотных остатков - "циклиновый бокс", отвечающий за связывание с циклинзависимой киназой (от англ. CDK - cyclin-dependent kinases). В клетках эукариотов существует примерно восемь различных CD К (CDK1-8), активирующихся различными циклинами (табл. 4-2).
Циклинзависимые киназы, связывая циклин, переходят в активную форму и могут фосфорилировать специфические белки, например факторы транскрипции, белки-ингибиторы факторов
156
транскрипции, которые регулируют синтез ферментов, обеспечивающих репликацию. Синтез каждого циклина начинается при подготовке к соответствующей фазе клеточного цикла, его концентрация в клетке повышается, а после окончания фазы резко падает до нуля. Завершившие свою работу комплексы циклинов и CDK связываются специфическими белками, ингибирующими их активность, и затем подвергаются разрушению.
Процесс, позволяющий живым организмам восстанавливать повреждения, возникающие в ДНК, называют репарацией. Все репарационные механизмы основаны на том, что ДНК - двухцепочечная молекула, т.е. в клетке есть 2 копии генетической информации. Если нуклеотидная последовательность одной из двух цепей оказывается повреждённой (изменённой), информацию можно восстановить, так как вторая (комплементарная) цепь сохранена.
Процесс репарации происходит в несколько этапов. На первом этапе выявляется нарушение комплементарности цепей ДНК. В ходе второго этапа некомплементарный нуклеотид или только основание устраняется, на третьем и четвёртом этапах идёт восстановление целостности цепи по принципу комплементарности. Однако в зависимости от типа повреждения количество этапов и ферментов, участвующих в его устранении, может быть разным.
Очень редко происходят повреждения, затрагивающие обе цепи ДНК, т.е. нарушения структуры нуклеотидов комплементарной пары. Такие повреждения в половых клетках не репарируются, так как для осуществления сложной репарации с участием гомологичной рекомбинации требуется наличие диплоидного набора хромосом.
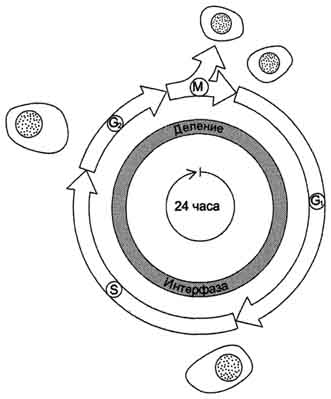
Рис. 4-20. Фазы клеточного цикла. После фазы М, в ходе которой происходит деление ядра (митоз) и цитоплазмы (цитокинез), дочерние клетки вступают в интерфазу нового цикла. Интерфаза начинается с фазы G1, в ходе которой активно происходят биосинтетические процессы, резко замедленные во время митоза. Фаза S - период синтеза ДНК; она заканчивается, когда содержание ДНК в ядре удвоится и хромосомы полностью реплицируются. Затем наступает фаза G2, в ходе которой происходят деление митохондрий и увеличение энергетических запасов клетки. Фаза G2продолжается до начала митоза, т.е. фазы М. В фазе М ядерная оболочка разрушается, формируются два новых ядра, цитоплазма делится с образованием двух дочерних клеток, имеющих по одному ядру. На рисунке представлен 24-часовой цикл.
А. Спонтанные повреждения
Нарушения комплементарности цепей ДНК могут происходить спонтанно, т.е. без участия каких-либо повреждающих факторов, например
Таблица 4-2. Циклимы и циклинзависимые киназы, регулирующие прохождение клеточного цикла
Циклин |
Киназа |
Функция |
D,E |
CDK4, CDK6 |
Регулирует переход клетки из G1-фазы в S-фазу |
А |
CDK2 |
Активирует синтез ДНК на начальной стадии S-фазы |
В |
CDK1 |
Регулирует переход клетки из G2-фазы в М-фазу |
157
в результате ошибок репликации, дезаминирования нуклеотидов, депуринизации.
Ошибки репликации
Точность репликации ДНК очень велика, но примерно один раз на 105-106 нуклеотидных остатков происходят ошибки спаривания, и тогда вместо пары нуклеотидов А-Т, G-С в дочернюю цепь ДНК оказываются включёнными нук-леотиды, некомплементарные нуклеотидам матричной цепи. Однако ДНК-полимеразы δ, ε способны после присоединения очередного нук-леотида в растущую цепь ДНК делать шаг назад (в направлении от 3'- к 5'- концу) и вырезать последний нуклеотид, если он некомплементарен нуклеотиду в матричной цепи ДНК. Этот процесс исправления ошибок спаривания (или коррекция) иногда не срабатывает, и тогда в ДНК по окончании репликации остаются некомплементарные пары, тем более, что ДНК-полимераза а лишена корректирующего механизма и "ошибается" чаще, чем другие полимеразы.
При неправильном спаривании в первичной структуре дочерней цепи ДНК необычные основания не появляются, нарушена только ком-плементарность. Система репарации некомплементарных пар должна происходить только на дочерней цепи и производить замену некомплементарных оснований только в ней. Ферменты, участвующие в удалении неправильной пары нуклеотидов, распознают матричную цепь по наличию метилированных остатков аденина в последовательностях -GATC-. Пока основания нуклеотидных остатков в дочерней цепи неметилированы, ферменты должны успеть выявить ошибку репликации и устранить её.
Распознавание и удаление (первый этап) некомплементарного нуклеотида происходят при участии специальных белков mut S, mut L, mut H. Каждый из белков выполняет свою специфическую функцию. Mut S находит неправильную пару и связывается с этим фрагментом. Mut Н присоединяется к метилированному (по аденину) участку -GATC-, расположенному вблизи некомплементарной пары. Связующим между mut S и mut Н служит белок mut L, его присоединение завершает образование активного фермента. Формирование комплекса mut S, mut L, mut Н на участке, содержащем ошибку, способствует проявлению у белка mut Н эндонуклеазной активности. Ферментативный комплекс гидролизует фосфоэфирную связь в неметилированной цепи (рис. 4-21).
К свободным концам цепи присоединяется экзонуклеаза (второй этап). Отщепляя по одному нуклеотиду в направлении от 3'- к 5'- концу дочерней цепи, она устраняет участок, содержащий некомплементарную пару. Брешь застраивает ДНК-полимераза β (третий этап), соединение основного и вновь синтезированного участков цепи катализирует фермент ДНК-лигаза (четвёртый этап). Для успешного функционирования экзонуклеазы, ДНК-полимеразы р и ДНК-лигазы необходимо участие в репарации хеликазы и SSB-белков.