

Антигенный дрейф вируса гриппа а
Годы отбора сывороток для исследования |
Титр антител к гемагглютининам вируса |
||
Н2 |
Н368 |
Н372 |
|
1968 |
1:100 |
<1:10 |
<1:10 |
1972 |
1:100 |
1:80 |
1:30 |
Примечание: Все сыворотки исследовали с тремя сероварами вируса. Серовар Н2 был идентичен вирусу, вызвавшему пандемию в 1957 г., Н368 – возбудителю 1968 г., Н372 – возбудителю эпидемии 1972 г.
Антигенный шифт (смена) обусловливает появление нового антигенного варианта (вида) вируса, не связанного либо отдаленно антигенно родственного ранее циркулирующим вариантам; то есть это радикальное резкое изменение одного или двух поверхностных антигенов. Подобное явление наблюдается достаточно редко (табл. 2). Антигенный шифт является результатом генетической рекомбинации между штаммами вирусов гриппа человека и животных. Это приводит к смене подтипа гемагглютинина или нейраминидазы, а иногда – обоих антигенов. В результате образуется принципиально новый антигенный вариант вируса (новый вид), обычно вызывающий пандемию, так как в популяции практически отсутствует иммунная прослойка; по мере образования последней процесс затухает.
Таблица 2
Основные антигенные шифты вируса гриппа а
Годы |
Серовар |
Название |
1947 |
H1N1 |
Испанский грипп |
1957 |
H2N2 |
Азиатский грипп |
1968 |
H3N2 |
Грипп Гонконг |
1976 |
H1N1 (свиной) |
Свиной грипп |
1977 |
H1N1 |
СССР (русский грипп) |
Первый штамм вируса гриппа, выделенный от человека, имел антигенную формулу Н0N1 (1933 год), а уже в 1947 г. был выделен серовар Н1N1; за последние 30 лет выделены серовары Н2N2, Н3N2.
Теории происхождения пандемических штаммов вируса гриппа. Все пандемии гриппа были вызваны вирусами гриппа А, претерпевшими шифт. Пандемия гриппа 1918 г. была вызвана вирусом с фенотипом Н1N1 (погибло около 20 миллионов человек); пандемия 1957 г. – вирусом Н2N2 (переболело более половины населения мира); 1968 г. – вирусом Н3N2. Для объяснения причин такой резкой смены типов вирусов гриппа А были предложены 2 гипотезы.
Согласно зооантропонозной гипотезе (авторы Вебстер и Тембол), вирус, вызвавший пандемию, после возникновения к нему иммунитета переходит на популяции млекопитающих или птиц. Затем в результате генетических рекомбинаций (чему способствует фрагментированный геном) между вирусами гриппа А человека и животных возникает рекомбинантный штамм с новым типом гемагглютинина, к которому у людей еще нет иммунитета, и он вызывает новую пандемию гриппа.
Согласно антропонозной гипотезе, выдвинутой Френсисом (США) и А.А. Смородинцевым (СССР), вирус, исчерпавший свои эпидемические возможности, не исчезает, а продолжает циркулировать в коллективе людей без заметных вспышек или длительно персистировать в организме человека. Через 10-20 лет, когда появится новое поколение людей, не имеющих к нему иммунитета, этот вирус возвращается и становится причиной новой пандемии. Данная гипотеза подтверждается тем, что вирус гриппа А с фенотипом Н1N1, исчезнувший в 1957 г., когда его вытеснил вирус Н2N2, вновь появился, после двадцатилетнего отсутствия, в 1977 г. Кроме того, по данным серологической археологии, пандемия 1889 г. была, очевидно, вызвана вирусом с фенотипом Н2N2, так как в 1957 г. антитела к нему были обнаружены только у людей старше 70 лет, не болевших во время пандемии 1957 г. Наконец, установлено, что грипп у людей вызывали и вызывают вирусы типа А только 3 или 4 фенотипов (Н1N1 (H0N1), H2N2, H3N2). Вместе с тем, выявилась новая эпидемиологическая особенность гриппа: если раньше каждый новый пандемический вариант полностью вытеснял своего предшественника, то с 1977 года вирусы с фенотипами Н1N1 и H3N2 как бы сосуществуют в коллективе, обладая, вероятно, до определенного времени равными эпидемическими возможностями. Антигенные дрейфы и шифты вирусов гриппа А являются главным препятствием при создании эффективных вакцин.
Культивирование вирусов гриппа А. Вирусы гриппа А культивируются в куриных эмбрионах и культурах клеток. В куриных эмбрионах вирусы гриппа А репродуцируются в течение 36-48 ч в амниотической и аллантоисной полостях при температуре 37˚С. Наиболее чувствительными к вирусу гриппа А являются первичные культуры клеток почек эмбриона человека и некоторых животных. Репродукция вируса в этих культурах сопровождается слабо выраженным цитопатическим действием, напоминающим спонтанную дегенерацию клеток.
Вирусы гриппа А можно культивировать в организме мышей, обезьян и приматов. Мыши в естественных условиях не инфицируются вирусами гриппа, но вирусы гриппа А могут быть адаптированы к мышам. Вирус размножается в верхних и нижних отделах респираторного тракта и после адаптации может вызвать у мышей пневмонию и смерть. Вид мышей А2G не чувствителен к гриппу и несет доминантную аллель (ген Мх), которая кодирует белок, индуцируемый интерфероном и ингибирует вирус гриппа.
Эпидемиология. Круг хозяев и распространение вируса. Вирусы гриппа А обычно вызывают заболевание у людей, свиней, лошадей и редко у птиц. С 1933 г. 3 серотипа гемагглютинина были идентифицированы у людей, 2 – у свиней и 2 – у лошадей. Все 16 серотипов были обнаружены у водоплавающих птиц, в основном у диких уток; они обычно не вызывают заболевание у этих хозяев, но служат резервуарами генетической информации гриппа А. Вирусы гриппа (птичьего происхождения) были случайно выделены от норок и китов.
Эволюция. Результаты анализа последовательностей каждого из генов вирусов гриппа А подтвердили предположение, что существует 5 специфических линий происхождения от хозяев, и что происхождение от птиц находится в эволюционном застое. Вирусы гриппа А, в настоящее время циркулирующие у людей, возможно, произошли приблизительно 150 лет назад от вирусов гриппа птиц. Предполагается, что катастрофическая «испанка» 1918 г. произошла от свиней (с подтверждением того, что свинья является промежуточным хозяином). Высоко вирулентные штаммы исчезли, но усилили текущее возникновение гриппа от человека. Вирусы гриппа А, циркулирующие у свиней и лошадей, также происходят из птичьего источника.
Э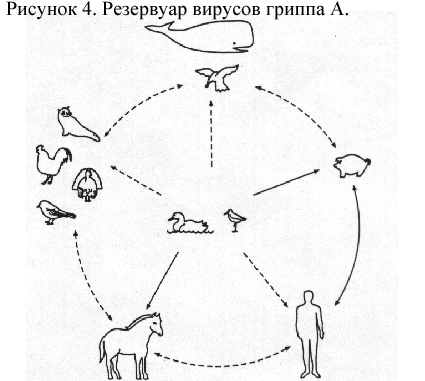 пидемиология.
Источником инфекции при гриппе являются
водоплавающие птицы семейств гусиных
и чайковых - исконный резервуар вирусов
гриппа для видов птиц и млекопитающих.
Вирусы гриппа птиц обычно распространяются
путем фекальной контаминации воды
(механизм передачи – фекально-оральный,
путь передачи – преимущественно водный,
в редких случаях – аэрогенно). Доказанной
является передача гриппа от свиней к
человеку и наоборот. Возможна передача
возбудителя от птиц свиньям и лошадям
и непрямая передача человеку от птиц
через свиней, а также передача возбудителя
от цыплят человеку. Среди людей источником
инфекции является больной гриппом
человек и вирусоноситель. Механизм
передачи – аэрогенный, реализующийся
преимущественно воздушно-капельным
путем. Заболевший человек становится
заразным за 24 ч до появления основных
симптомов заболевания и представляет
эпидемиологическую опасность в течение
48 ч после их исчезновения.
пидемиология.
Источником инфекции при гриппе являются
водоплавающие птицы семейств гусиных
и чайковых - исконный резервуар вирусов
гриппа для видов птиц и млекопитающих.
Вирусы гриппа птиц обычно распространяются
путем фекальной контаминации воды
(механизм передачи – фекально-оральный,
путь передачи – преимущественно водный,
в редких случаях – аэрогенно). Доказанной
является передача гриппа от свиней к
человеку и наоборот. Возможна передача
возбудителя от птиц свиньям и лошадям
и непрямая передача человеку от птиц
через свиней, а также передача возбудителя
от цыплят человеку. Среди людей источником
инфекции является больной гриппом
человек и вирусоноситель. Механизм
передачи – аэрогенный, реализующийся
преимущественно воздушно-капельным
путем. Заболевший человек становится
заразным за 24 ч до появления основных
симптомов заболевания и представляет
эпидемиологическую опасность в течение
48 ч после их исчезновения.
Патогенез гриппа включает все 7 стадий, характерных для патогенеза циклических вирусных заболеваний (глава «Вирусный инфекционный процесс»).
Вирусы гриппа А посредством гемагглютинина прикрепляются к плоскому и мерцательному эпителию верхнего отдела дыхательных путей. В эпителиальных клетках происходит первичная репродукция вирусов. Размножение протекает с исключительно высокой скоростью, что достигается благодаря фрагментированному геному вирусов гриппа А и объясняет короткий инкубационный период – 1-2 дня. Быстроте репродукции вирусов способствует распространение многих сотен вирионов, подготовленных лишь одной зараженной клеткой. В дальнейшем вирусы попадают в кровь и разносятся по всему организму. Под действием вирусов активируется система протеолиза и повреждается эндотелий капилляров, что приводит к повышенной проницаемости сосудов, кровоизлияниям и дополнительному повреждению тканей различных органов (трахея, бронхи, миокард, легкие, головной мозг, почки). Вирусы гриппа А, попадая в кровь, вызывают угнетение кроветворения и иммунной системы, развивается лейкопения и гиперсупрессорный вариант иммунодефицита. Поражение мерцательного эпителия дыхательных путей сопровождается его разрушением, что является входными воротами для проникновения бактерий в легкие. Появляется опасность развития бактериальной суперинфекции – бронхита, пневмонии. Белок NS1 вируса гриппа А способен индуцировать апоптоз в чувствительных клетках.
Иммунитет обеспечивается системой интерферонов, натуральными киллерами, Т-киллерами и специфическими антителами. Интерфероны (преимущественно α-интерфероны) ингибируют репродукцию вируса в эпителиальных клетках, а также стимулируют функциональную активность натуральных киллеров. Последние разрушают вирусинфицированные клетки, что способствует элиминации возбудителя из организма. Индукторами продукции интерферонов являются сами вирусы гриппа, однако продукция интерферонов существенно угнетается при высоких инфицирующих дозах вируса.
Антигенспецифические Т-киллеры разрушают вирусинфицированные эпителиоциты, предоставляя тем самым доступ к вирусам гриппа иммуноглобулинам. Последние, взаимодействуя с антигенными детерминантами вирусов, образуют ЦИК. При низкой аффинности иммуноглобулинов инактивации вируса в ЦИК не происходит, что может вызвать инфицирование здоровых эпителиоцитов. Элиминация ЦИК из организма больного гриппом осуществляется системой макрофагов. Завершенность фагоцитоза ЦИК зависит от их молекулярности: наиболее интенсивно элиминируются крупномолекулярные ЦИК, тогда как средние и мелкие ЦИК могут длительно циркулировать во внутренней среде организма, оседая в паренхиматозных органах (почках, легких, мозге), а также сосудах микроциркуляторного русла, что вызывает дополнительное их поражение.
Секреторные иммуноглобулины А, выводимые на поверхность слизистых оболочек верхних дыхательных путей, вызывают инактивацию вирусов гриппа и способствуют их фагоцитозу. Основными защитными антителами при гриппе являются секреторные Ig А и сывороточные Ig М и G к молекуле гемагглютинина, которые нейтрализуют инфективность вирусов и отвечают за формирование стойкости к инфекции. Гуморальный ответ на гемагглютинин является специфическим для данного вида вирусов, однако антигенный дрейф позволяет вирусам избежать инактивации антителами. Антитела к нейраминидазе не предотвращают инфекцию, но уменьшают распространение вирусов в организме. Иммунитет при гриппе является родо- и видоспецифическим, сохраняющимся в течение многих десятилетий.
Клиника. Грипп начинается остро с озноба, лихорадки (39-40˚С), головной боли, слабости, ломоты в костях и суставах, заложенности носа со скудным отделяемым, непродуктивного кашля. Более тяжелое течение гриппа может развиться, если возникнет первичная гриппозная пневмония или вторичная бактериальная пневмония. Длительность болезни у взрослых – в среднем 7 дней. Дети, которые болеют гриппом первый раз в жизни, могут содержать в себе вирус в течение 13 дней.
Лабораторная диагностика гриппа включает вирусоскопический, вирусологический и серологический методы диагностики. Материалом для исследования являются мазки, секрет и смывы из носоглотки, кровь, спинномозговая жидкость и секционный материал.
Экспресс-диагностика. Используется метод иммунофлюоресценции антител (прямой метод Кумбса). Позволяет провести исследование в течение 2-3 ч от момента взятия материала. Клетки цилиндрического эпителия слизистой оболочки нижней носовой раковины и задней стенки глотки отбирают сухими ватными тампонами и помещают в среду для транспортировки вирусов. В лаборатории тампоны отжимают, а взвесь центрифугируют. Из осадка клеток готовят мазки на предметных стеклах. При посмертном выявлении антигенов вируса гриппа делают отпечатки кусочков ткани легких, а также готовят препараты со слизистой оболочки трахеи и бронхов, соскабливая клетки эпителия. Препараты обрабатываются противогриппозными иммуноглобулинами, нагруженными флюорохромами, инкубируются в течение часа, после чего отмываются физиологическим раствором. Вирусинфицированные клетки при специфическом взаимодействии с иммуноглобулинами и последующем их рассмотрении в люминесцентном микроскопе проявляют свечение. Локализация и характер свечения зависят от стадии развития вируса гриппа в клетках, а также от срока возникновения гриппозной инфекции. В первые дни болезни антиген чаще локализуется в ядрах клеток цилиндрического эпителия при одновременном диффузном или гранулярном свечении цитоплазмы в тех же самых или других клетках. Часто выявляется равномерное гомогенное свечение всей клетки. В случае затухания инфекции свечение чаще всего наблюдается в цитоплазме или в ее части в виде отдельных гранул. Путем сопоставления количества пораженных клеток, находящихся в поле зрения, со сроком заболевания, отмечают наибольшее их количество (4-10) в первые дни болезни, чем в последующие. Диагностическим является специфическое свечение 5 и больше клеток цилиндрического эпителия с яркостью не менее «++».
Реакция непрямой гемадсорбции основывается на способности клеток цилиндрического эпителия верхних дыхательных путей, пораженных вирусом гриппа, адсорбировать на своей поверхности эритроциты, сенсибилизированные противогриппозными антителами. Используется 0,25 % взвесь сенсибилизированных эритроцитов барана. После 30 минут экспозиции при комнатной температуре взвесью клеток заполняют счетную камеру Горяева и микроскопируют в световом микроскопе с помощью фазово-контрастной оптики. Если в исследуемом препарате выявляют 4-5 клеток цилиндрического эпителия с адсорбированными на них двумя и более эритроцитами, реакцию считают положительной.
Реакция пассивной гемагглютинации. Антиген вируса гриппа в исследуемом материала выявляют с помощью эритроцитарного антительного диагностикума, который соединяют с освобожденным от слизи и гетерогемагглютининов смывом в соотношении 1:20. При наступлении гемагглютинации реакцию считают положительной. В качестве экспресс-метода диагностики может быть использован ИФА.
Вирусологический метод. Исследуемым материалом являются смывы из носоглотки, секционный материал, спинномозговая жидкость. Вирусы выделяют на куриных эмбрионах и в культурах клеток.
Выделение вирусов гриппа на куриных эмбрионах является наиболее доступным. 10-11-дневные куриные эмбрионы заражают инфекционным материалом в объеме 0,1 мл в амниотическую или 0,2 мл – в аллантоисную полости, после чего эмбрионы выдерживают при 33-34˚С в течение 72 ч (оптимальные условия репродукции вирусов А и В). Полученный из куриных эмбрионов вируссодержащий материал исследуют на присутствие феномена гемагглютинации в реакции гемадсорбции (РГА) с эритроцитами кур или морских свинок. Если результаты РГА отрицательные, после пассажей исследование материала заканчивают. В случае наличия агглютинации эритроцитов проводят титрование гемагглютинирующего вируса в РГА.
Каждый вирусный антиген титруют в 2 параллельных рядах лунок планшета в разведениях от 1:10 до 1:2560. Разница титра в 2 рядах не должна превышать одного двукратного разведения. Если она больше, титрование следует повторить. Обсчитывая титр (в случае двукратной разницы), рассчитывают его среднеарифметическое значение. После определения гемагглютинирующего титра свежевыделенного вируса гриппа готовят его рабочее разведение, содержащее 4 ГАЕ (гемагглютинирующие единицы) в заданном объеме. Идентифицируют изолированный вирус гриппа в РТГА (реакция торможения гемагглютинации), используя коммерческие диагностические сыворотки против возбудителей гриппа А1 (Н1N1), А2 (Н2N2), А3 (Н3N2), В и С. Кроме того, тип вируса гриппа можно определить в реакции связывания комплемента (РСК).
Выделение вирусов гриппа в культурах клеток. Используются однослойные трипсинизированные культуры клеток эмбриона человека и однодневных цыплят. ЦПД в случае заражения вирусами гриппа характеризуется дегенерацией слоя клеток. ЦПД появляется с 3 по 10 день с момента заражения. Вирусы гриппа А в культуре клеток развиваются медленнее, их ЦПД выражается в появлении фестончатых клеток или клеток с вакуолизацией цитоплазмы, которые слущиваются в процессе дегенерации. Гемадсорбция выявляется значительно раньше, чем ЦПД. Если титр гемагглютининов в культуральной жидкости составляет 1:8 и больше, выделенные на культуре клеток вирусы гриппа идентифицируют в РТГА, если титр меньше, чем 1:8, собранный материал культивируют для повышения титра, либо проводят идентификацию в реакции торможения гемадсорбции (РТГАдс) на инфицированной культуре. Для торможения гемадсорбции титр иммунной сыворотки должен быть не менее чем 1:160 и соответствовать антигенной структуре выделенного вируса. Тип гемагглютинина вируса гриппа при вирусологическом методе исследования устанавливают в РТГА, подтип нейраминидазы – в реакции ингибирования нейраминидазной активности.
Серологический метод основывается на выявлении увеличения титра противогриппозных антител в динамике заболевания. Для серологической диагностики гриппа используют РНГА, РСК, РТГА, причем последние 2 реакции – наиболее часто. В процессе серологического исследования сывороток крови в РТГА используют последовательные двукратные разведения их на изотоническом растворе натрия хлорида, гриппозный антиген (4 ГАЕ) и взвесь эритроцитов кур или человека с группой крови О (I). Для серологической диагностики гриппа РСК осуществляют в тех же самых условиях, что и для идентификации изолированных вирусов. В РНГА антитела выявляют с помощью стандартных лиофилизированных антигенных эритроцитарных диагностикумов.
Лечение. Для лечения гриппа используют α-интерфероны, обладающие наибольшим противовирусным действием (реаферон – человеческий генно-инженерный рекомбинантный α2-интерферон, который назначают от 500 000 до 1 000 000 ЕД 3 раза в день внутримышечно на протяжении 5-7 дней). Противовирусным действием обладают также индукторы продукции эндогенного интерферона (мефенаминовая кислота, амизон). Выраженное противовирусное действие оказывают химиопрепараты ремантадин, амантадин. Для лечения гриппа применяют также противогриппозные иммуноглобулины. При осложненном течении гриппозной инфекции показаны антибиотики широкого спектра действия.
Профилактика. Неспецифическая профилактика гриппа включает ранее выявление, изоляцию и санацию источника инфекции (больного человека), а также разрыв механизма и пути передачи вирусов гриппа. С этой целью устанавливается противоэпидемический режим в организованных коллективах (разобщение лиц, ношение масок, соблюдение противоэпидемического режима в больничных отделениях). В очагах инфекции проводится ультрафиолетовое облучение и влажная уборка помещений с использованием дезинфицирующих средств. Массовая неспецифическая профилактика включает использование индукторов продукции эндогенного интерферона (амизон, мефенаминовая кислота).
Специфическая профилактика включает в себя иммуноглобулинопрофилактику и вакцинопрофилактику. Используются донорские противогриппозные иммуноглобулины в соответствии с возрастными дозами. Для вакцинопрофилактики применяются следующие типы противогриппозных вакцин: 1) живая аттенуированная гриппозная вакцина (аллантоисная и культуральная); 2) убитая цельновирионная гриппозная вакцина; 3) субвирионная гриппозная вакцина; 4) субъединичная гриппозная вакцина, содержащая только гемагглютинин и нейраминидазу.
Наиболее эффективными являются субъединичные гриппозные вакцины, среди которых имеется моновакцина (Н1N1), дивакцина (Н1N1+Н3N2) и тривакцина (А/H1N1+A/H3N2+B). В настоящее время в Украине широко используются зарубежные гриппозные вакцины «Флюарикс» (Бельгия), «Ваксигрипп» (Франция) и «Инфлувак» (Голландия). Эти вакцины являются субвирионными гриппозными тривакцинами. Одна доза вакцины (0,5 мл) вводится внутримышечно или подкожно, иммунитет развивается в течение 3-4 недель. Вакцинацию осуществляют в периоды наибольшего риска развития эпидемий. Вакцинация наиболее показана лицам младшего и преклонного возрастов, а также сотрудникам лечебно-профилактических учреждений. Вакцинацию следует проводить за 3-4 недели до начала эпидемии.
§ 3. Род Influenzavirus В.
Род включает вирусы гриппа В человека, сходные с вирусами гриппа А, но не идентичные им.
Вирус гриппа В
История открытия. Вирусы гриппа В открыты в 1940 г. в США Т. Френсисом и Р. Меджиллом.
Особенности вируса. Вирионы вируса гриппа В имеют диаметр 90-100 нм и строение, типичное для ортомиксовирусов. Атипичные нитевидные формы вирионов встречаются редко. Суперкапсид содержит гемагглютинин, нейраминидазу и белки ионных каналов, представленные NВ-протеином, который составляет внутреннюю поверхность суперкапсида. Гемагглютинин является основным поверхностным антигеном. Капсид построен по спиральному типу симметрии из М1-белка.
Геном представлен 8 сегментами негативной РНК длиной 14,6*103 нуклеотидов. Для сегментов РНК вирусов гриппа В конформация в виде замкнутого кольца не доказана. Сегменты 6 и 8 кодируют по 2 белка, остальные – по 1. Каждый сегмент РНК связан с NP, PB1, PB2 и PA-белками. В вирионе содержится специфический для вируса гриппа В белок ВМ2, функция которого неизвестна.
Культивирование. Вирусы гриппа В в куриных эмбрионах размножаются медленнее, чем вирусы гриппа А (72 ч против 48 ч), и при более низких температурах (33-35˚С).
Антигенная структура. Внутренние белки М1 и NР являются родоспецифичными; гемагглютинин и нейраминидаза – видоспецифичными. Перекрестного иммунитета между вирусами гриппа А и В нет. По антигенным свойствам внутри единственного вида вирусов гриппа В известно 5 подвидов. Антигенная изменчивость вирусов гриппа В менее выражена, чем у вирусов гриппа А.
Эпидемиология. Источником инфекции при гриппе В является больной человек или вирусоноситель. Механизм передачи инфекции – аэрогенный, реализуемый воздушно-капельным путем. Вирусы гриппа В не вызывают пандемий и обычно являются причиной локальных вспышек и эпидемий, иногда охватывающих одну или несколько стран. Вспышки гриппа В могут совпадать со вспышками гриппа А или предшествовать им. Вирусы гриппа В циркулируют преимущественно в человеческой популяции, чаще вызывая заболевание у детей. Вирусы гриппа В выделены от тюленей Северного моря, которые были инфицированы вирусами, вызвавшими до этого заболевания у людей.
Патогенез гриппа В сходен с таковым при гриппе А.
Клиника. Симптомы заболевания при гриппе В сходны с клиническими проявлениями гриппа А. Дифференциальный диагноз осуществляется лабораторными методами.
Лабораторная диагностика гриппа В включает использование вирусологического, серологического и вирусоскопического методов. При вирусологическом исследовании идентификация вирусов проводится в РТГА с использованием иммунной сыворотки против вирусов гриппа В. Серологическая диагностика осуществляется в РПГА с В-гриппозным диагностикумом. Вирусоскопический метод включает применение люминесцентной и иммунной электронной микроскопии.
Лечение. Показаны ремантадин, интерфероны и их индукторы.
Профилактика. Неспецифическая профилактика аналогична таковой для гриппа А. Специфическая профилактика проводится вакцинами «Флюарикс», «Ваксигрипп» и «Инфлувак», содержащими, наряду с гемагглютининами и нейраминидазами двух видов вируса гриппа А, компоненты вируса гриппа В/Бейджинг/184/93, или В/Шенгдонг/7/97. Защитный титр антител в РПГА после вакцинации – 1:40 и более.
§ 4. Род Influenzavirus С.
Род представлен вирусом гриппа С, вызывающим грипп у человека.
Вирус гриппа С
История открытия. Впервые выделен Р. Тейлором в 1947 г. из носоглоточного смыва больного гриппом посредством заражения куриных эмбрионов.
Особенности
вируса.
Вирионы вируса гриппа С сферические,
диаметром 100-120 нм. Встречаются нитевидные
формы (рис. 5). Строение вириона типичное
для семейства ортомиксовирусов.
Суперкапсид вируса гриппа С имеет только
1 тип гликопротеина, который называется
гемагглютинин-эстераза. Этот гликопротеин
отвечает за связывание с рецепторами
клетки, проникновение в клетку и
разрушение рецепторов при в ыделении
вирусов. Кроме гликопротеина, на
поверхности суперкапсида имеется
мембранный белок СМ2, выступающий как
ионный канал для проведения протонов
внутрь вириона, что необходимо для
«раздевания» вируса. СМ2-белок выстилает
также внутреннюю поверхность суперкапсида.
Нейраминидаза на суперкапсиде отсутствует.
Капсид вируса гриппа С спирального типа
симметрии и образован белком М1. Геном
представлен 7 сегментами негативной
РНК; седьмой сегмент кодирует 2 белка,
остальные – по одному. Длина РНК
составляет 12,9*103
нуклеотидов. Конфигурация сегментов
РНК в виде замкнутого кольца для вирусов
гриппа С не доказана. С сегментами РНК
связаны NP,
PB1, PB2 и PA-белки.
Геном вируса кодирует также NS2-белок,
обеспечивающий выход РНК из ядра клетки.
ыделении
вирусов. Кроме гликопротеина, на
поверхности суперкапсида имеется
мембранный белок СМ2, выступающий как
ионный канал для проведения протонов
внутрь вириона, что необходимо для
«раздевания» вируса. СМ2-белок выстилает
также внутреннюю поверхность суперкапсида.
Нейраминидаза на суперкапсиде отсутствует.
Капсид вируса гриппа С спирального типа
симметрии и образован белком М1. Геном
представлен 7 сегментами негативной
РНК; седьмой сегмент кодирует 2 белка,
остальные – по одному. Длина РНК
составляет 12,9*103
нуклеотидов. Конфигурация сегментов
РНК в виде замкнутого кольца для вирусов
гриппа С не доказана. С сегментами РНК
связаны NP,
PB1, PB2 и PA-белки.
Геном вируса кодирует также NS2-белок,
обеспечивающий выход РНК из ядра клетки.
Антигенная структура. Белки М1 и NP являются родоспецифичными. Антигенный шифт и дрейф у вирусов гриппа С отсутствует.
Культивирование. Характерна низкая адаптационная способность вируса гриппа С к клеткам куриных эмбрионов и различным клеточным культурам. В амнионе куриного эмбриона вирусы развиваются при 33˚С в течение 72-96 ч инкубирования. Вирусы гриппа С удовлетворительно растут в клетках почек обезьян. В других клеточных линиях репродукция вирусов гриппа С либо вообще не регистрируется, либо отмечается формирование неполных вирионов, обладающих гемагглютинирующей, но не инфекционной активностью. Вирусы плохо агглютинируют эритроциты морской свинки, крыс и кур, в связи с чем серологические реакции гемагглютинации проводят при низкой температуре (4-6˚С).
Эпидемиология. Источник инфекции – больной человек или вирусоноситель. Механизм передачи – аэрогенный, реализуемый воздушно-капельным путем. Грипп С часто совпадает с эпидемией гриппа А.
Патогенез. Этапы патогенеза аналогичны таковым для патогенеза гриппа А и В. Вследствие более низкой патогенности вирусов гриппа С поражение тканей менее значительно, чем при гриппе А и В.
Клиника. Чаще инфицируются дети. Симптомы болезни обычно очень легкие либо не проявляются вообще. В ряде случаев клиническая картина такая же, как при легких и среднетяжелых формах гриппа А. Дифференциальная диагностика гриппа С осуществляется лабораторными методами.
Лабораторная диагностика. Основной диагностический метод – серологический (РПГА с эритроцитарным диагностикумом, содержащим антигены вирусов гриппа С). Реже применяется вирусологический и вирусоскопический методы.
Лечение. Показаны ремантадин, интерфероны и их индукторы.
Профилактика. Неспецифическая профилактика аналогична профилактике гриппа А и В. Вакцины против гриппа С не разработаны.
§ 5. Род Thogotovirus.
Тоготовирусы являются передаваемыми клещами ортомиксовирусами, инфицирующими людей и домашний скот в Африке, Азии и Европе. Вирусы Тогото были впервые изолированы в лесу Тогото около Найроби (Кения). Инфицируют крыс, буйволов, верблюдов, ослов, крупный рогатый скот, овец и людей. У людей тоготовирусы были изолированы из спинномозговой жидкости при билатеральном оптическом нейрите и из крови при менингите.
Сходство тоговирусов с вирусами гриппа основано на общих молекулярных и морфогенетических свойствах. Геном вируса представлен 6 сегментами однониточной «-»-РНК, один из сегментов кодирует белок, сходный с РА-белком вирусов гриппа. Концы сегментов сходны с концами вирусов гриппа. При репликации тоготовирусы проходят ядерную фазу, но синтез вирусной мРНК не сопровождается захватом кэпов мРНК хозяина, как это происходит у вирусов гриппа.