

МОСКОВСКИЙ ГОСУДАРСТВЕННЫЙ УНИВЕРСИТЕТ
имени М.В. ЛОМОНОСОВА
Биологический факультет
Методическая разработка по спецкурсу
ГАМЕТОГЕНЕЗ
Москва, 2013 г.
Мейоз в половых клетках
Общая характеристика
Профаза 1 мейоза состоит из 5 последовательных стадий: лептотены, зиготены, пахитены, диплотены и диакинеза.
По завершении последнего гониального деления в ядрах происходит дополнительный синтез ДНК, в результате чего вступающие в мейоз клетки содержат удвоенное по сравнению с нормальным количество ДНК (2n4c). При этом каждая хромосома состоит из 2-х сестринских хроматид, прикрепленных к общей центромере.
В профазу 1 вступают прелептотенные гонии, закончившие стадию размножения.
На стадии лептотены (стадия тонких нитей) хромосомы становятся видимыми в виде тонких нитей. Хроматин хроматид оформляется в очень тонкие нити, не позволяющие в этом клубке идентифицировать отдельные хромосомы. Так как репликация уже произошла, то каждая хромосома представлена двумя параллельно лежащими хроматидами.
На стадии зиготены (стадия сопряженных нитей) гомологичные хромосомы расположены парами (конъюгируют), образуя биваленты. Гомологичные хромосомы располагаются одна вдоль другой. Такое спаривание называют синапсисом – характерная черта мейоза, которой нет у митоза. Между спаренными хромосомами образуется синаптонемальный комплекс (рис. 1). Он представляет собой структуру, напоминающую лестницу, с центральным элементом и двумя полосами по бокам. Поскольку хроматин связан с этими боковыми полосами, хроматиды оказываются соединенными. Спаренные хромосомы связаны с ядерной мембраной. Считается, что она способствует удержанию вместе гомологичных хромосом.

Рис.1 Синаптонемальный комплекс. А. Гомологичные хромосомы, удерживаемые вместе на стадии пахитены мейоза в ооците Neoteeiella. Б. Схема, поясняющая строение синаптонемального комплекса
СК имеет ширину 0,2 мкм, и сотоит из трех параллельных темных лент, закрученных вдоль оси бивалента на протяжении всей его длины. Два наружных элетронно-плотных слоя этой трехчленной структуры называются боковыми элементами (их ширина до 600 А), средняя продольная полоса – центральный элемент (ширина до 400 А) лежит в светлом пространстве между боковыми элементами. Центральное пространство (ширина до 1200 А) содержит поперечные волокна, соединяющие боковые и центральный элементы. Боковые и центральный элементы могут отличаться по ультраструктуре. Боковые элементы состоят из белка класса основных гистоновых белков. Центральный элемент также состоит из белков. СК похож на застежку «молнию»: каждый боковой элемент состоит из наружной поверхности, образованной выстроенными в ряд синаптомерами (из ДНК), и внутренней поверхности, образованной выстроенными в ряд зигосомами (белок). Центральный элемент состоит из упорядоченно расположенных хвостиков зигосом, и он также содержит белок, но не ДНК. Конфигурация, образованная четырьмя хроматидами и синаптонемальным комплексом, называется бивалентом (у человека - 23).
На стадии пахитены (стадия толстых нитей) хроматиды утолщаются и укорачиваются, в световом микроскопе можно наблюдать отдельные хроматиды. Происходит кроссинговер (перекрест хромосом), во время которого хроматиды спаренных хромосом обмениваются генетическим материалом (гомологичными генами).
Кроссинговер продолжается и на следующей стадии – диплотене (стадия двойных нитей). Синаптонемальный комплекс разрушается и обе гомологичные хромосомы начинают обособляться, между ними остаются места сцепления – хиазмы (здесь происходит кроссинговер, рис. 2).
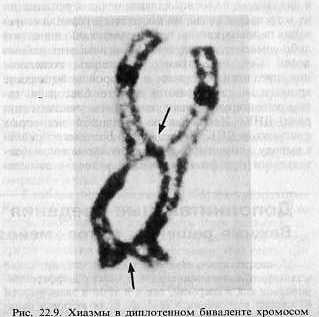
Рис.2 Хиазмы в диплотенном биваленте хромосом (обозначены стрелками).
На стадии диплотены высок уровень транскрипции генов. Как у мужских, так и у женских половых клеток хромосомы принимают вид «ламповых щеток», активно синтезируется РНК.
Диакинез (стадия обособления двойных нитей): центромеры расходятся, так что хромосомы остаются соединенными только в областях хиазм и концов хроматид. В конце диакинеза происходит разрушение ядерной оболочки и миграция хромосом к метафазной пластинке.
Первое деление мейоза – редукционное, в дочерние клетки расходятся гомологичные хромосомы, ядерная формула дочерних клеток 1n2c. В результате неправильного расхождения хромосом, в половых клетках могут либо появляться дополнительные хромосомы, либо какая-либо хромосома может отсутствовать вовсе.
После короткого интеркинеза (без синтетической фазы) наступает второе деление мейоза – эквационное. Каждая из образовавшихся клеток получает одну из двух хроматид. В результате кроссинговера образовавшиеся после мейоза каждая из 4-х дочерних клеток имеет свой, отличный от других и от дочерней клетки, набор хромосом. У человека в результате мейоза может сформироваться 223 разнообразных типов гаплоидных клеток, и это число увеличивается за счет вклада кроссинговера.
Особенности мейоза в ходе оогенеза
Блок мейоза: на стадии диплотены-диакинеза клетка может задерживаться долгие годы.
Усиленный синтез РНК на стадии диплотены (период роста), запасание РНП-частиц.
Неравномерность мейотических делений: во время телофазы полярное тельце практически не получает цитоплазмы, но получает ровно половину генетического материала, а яйцеклетка – вторую половину генетического материала и практически всю цитоплазму с запасенными веществами. В результате мейоза из одного ооцита I порядка образуется 1 зрелая яйцеклетка и 2 или 3 полярных тельца. Полярные тельца вскоре погибают или (у некоторых насекомых) сливаются с яйцеклеткой, участвуя в партеногенезе.
Преобразование (дифференцировка) проходит до мейотических делений в период роста (запасание желтка на стадии большого роста).
Особенности мейоза в ходе сперматогенеза
Главными и общими для процесса спермиогенеза у всех животных являются следующие моменты:
изменение формы и объема ядра, конденсация хроматина, коррелирующая с его метаболической инертностью;
образование акросомы – органеллы, обеспечивающей размягчение и растворение яйцевых оболочек при оплодотворении;
образование аппарата движения – жгутика или жгутиков или системы микротрубочек, заменяющей жгутик;
установление связи с питающими клетками;
сбрасывание цитоплазмы и органелл (рибосом, ЭПР, АГ и т.д.) и индивидуализация сперматозоидов (освобождение от цитоплазматических мостиков).
Процесс спермиогенеза можно условно разделить на несколько стадий:
фаза интенсивного развития АГ в цитоплазме сперматиды. Сперматида имеет округлую форму, сферическое ядро с диффузным хроматином. На этой стадии начинают формироваться проакросомные гранулы, содержащие литические ферменты (кислую фосфатазу, гиалуронидазу, неспецифическую этеразу, арилсульфатазу и др.) и являющиеся производными АГ. Митохондрии вначале дисперсно расположены в цитоплазме, затем занимают положение непосредственно под цитоплазматической мембраной, парные центриоли мигрируют к полюсу ядра, противоположному полюсу, занятому акросомным пузырьком.
Фаза «шапочки», во время которой на переднем конце ядер сперматозоидов выявляется акросома – небольшое электронноплотное полушарие у переднего конца ядра. При формировании акросомы несколько проакросомных гранул сливаются в одну большую гранулу, которая контактирует с ядерной оболочкой. В этот период гликопротеины переносятся из АГ в акросому с помощью мембранных пузырьков.
Фаза формирования удлиненной сперматиды – ядро сперматозоида удлиняется, начинается конденсация хроматина. В клетке хорошо выявляется ось симметрии, проодящая через передний и задний полюсы ядра. Акросома, расположенная на переднем полюсе ядра, по мере удлинения ядра приходит в тесный контакт с цитоплазматической мембраной, формируется субакросомальное пространство. АГ отделяется от акросомы. Пара центриолей, расположенных на периферии цитоплазмы, передвигается к хвостовому полюсу ядра (ранее они играли роль клеточного центра при построении веретен деления). Одна из них, вытянутая вдоль длинной оси будущего жгутика (дистальная центриоль), дает начало аксонеме жгутика. Другая центриоль, ориентированная перпендикулярно аксонеме, сохраняется и носит название проксимальной. Аксонема отходит от центриоли, доходит до цитоплазматической мембраны ранней сперматиды и вытягивается, образуя выступ. Митохондрии образуют спиральный воротникообразный футляр вокруг средней части будущего жгутика.
Фаза созревания, во время которой происходит удаление цитоплазматической капли с шейки сперматозоида в виде остаточных телец (резидуальных капель) и фагоцитоз их клетками Сертоли или эпителиальными клетками эпидидимиса. В цитоплазме около задней части ядра и у основания жгутика образуется манжетка из микротрубочек, которые идут параллельно поверхности ядра до апикального полюса и у некоторых видов окружают акросому. Это- временные органеллы, они не встречаются в зрелых сперматозоидах. Они участвуют в поддержании вытянутой формы ядра, сегрегации нуклеоплазмы.
Теория клонального развития мужских половых клеток
Незавершенный цитокинез во время митотических и мейотических делений мужских половых клеток приводит к тому, что количество клеток в синцитиях увеличивается в тысячи раз к тому времени, когда сперматозоиды выходят в просвет семенных канальцев. Каждая клеточная ассоциация состоит из нескольких синцитиев, находящихся на разных стадиях дифференцировки. В семеннике (человека) каждая клеточная ассоциация занимает четко очерченный сектор эпителия извитого семенного канальца.
Характерной и уникальной особенностью сперматогенеза является то, что в процессе митотических и мейотических делений не происходит полного разделения цитоплазмы образующихся дочерних клеток (цитокинеза). Все дифференцирующиеся сперматозоиды, происходящие из одной стволовой сперматогенной клетки, остаются связанными цитоплазматическими мостиками (фузомами) вплоть до выхода в просвет семенного канальца и образуют синцитий. Полагают, что наличие цитоплазматических мостиков обеспечивает высокую синхронность дифференциации мужских половых клеток, входящих в состав одного клона. Таким образом, развитие гаплоидных сперматозоидов регулируется полиплоидным геномом, что особенно важно в отношении мРНК половых хромосом, так как многие белки, важные для спермиогенеза контролируются генами X- и Y-хромосом.
Существует и другая точка зрения, согласно которой белки, регулирующие течение поздних стадий сперматогенеза (спермиогенеза), накапливаются в период существования в клетке диплоидного генома и продолжают работать в то время, когда геном становится гаплоидным.
Морфология зрелого сперматозоида
Сперматозоид человека
Зрелые нормальные сперматозоиды состоят из головки, шейки и жгутика, в длину сперматозоид человека 0,05 мм. Плоская головка имеет уплощенную «ложковидную» форму с заостренным передним полюсом, диаметр головки – всего 5 мкм. Передняя часть головки сперматозоида покрыта акросомой.
Большую часть головки зрелого сперматозоида составляет ядро с плохо различимой ядерной оболочкой и гомогенной, плотно упакованной массой хроматина. Примерно 2/3 передней поверхности ядра покрыто акросомой, заполненной гомогенным веществом средней электронной плотности. Акросома занимает 25-60% ядерной поверхности, содержит ферменты спермолизины.
Достаточно часто в ядрах нормальных сперматозоидов выявляются небольшие вакуоли, которые считаются остаточными явлениями конденсации хроматина. Вакуоли могут содержать гранулярный или филаментарный материал, липидные или другие включения.
Шейка сперматозоида имеет сложное строение. На мазке она выявляется как утолщение жгутика. У сформированного сперматозоида шейка представляет собой усеченный конус, содержащий центриолярный аппарат. Последний состоит из перпендикулярно расположенных проксимальной и дистальной центриолей. От дистальной центриоли отходит аксонема, составляющая морфологическую основу жгутика. Аксонема жгутика организована по универсальной схеме, характерной для большинства жгутиков эукариотических клеток. Она состоит из 9 периферических пра микротрубочек, расположенных по окружности, и центральной пары микротрубочек (9х2)+2. каждый периферичесий дуплет состоит из полной микротрубочки (субъединицы А) и примыкающей к ней неполной микротрубочки (субъединицы В). От субъединицы А каждого дуплета в сторону субъединицы В следующего дуплета отходят выросты, состоящие из белка динеина, проявляющего Mg-зависимую АТФ-азную активность. Эти выросты называются наружными и внутренними динеиновыми ручками. Дуплеты соединены тонкими нексиновыми мостиками, радиальные «спицы» прикреплены к листку, окружающему центральные микротрубочки. Вся эта сложная структура обеспечивает скользящее движение микротрубочек, лежащее в основе ундуляции жгутика. Микротрубочки аксонемы прикреплены к проксимальной центриоли непосредственно под головкой сперматозоида и тянутся от шейки до кончика жгутика сперматозоида. Вокруг аксонемы расположены добавочные структуры. В зависимости от их строения, морфологически жгутик можно разделить на среднюю часть, основной участок и концевой участок. В средней части аксонема окружена дополнительным слоем из 9 наружных плотных фибрилл и митохондрий, в основном участке они заменяются фиброзным слоем, в концевом участке остается только аксонема. В средней зоне жгутика нормальных сперматозоидов митохондрии образуют спираль вокруг аксонемы, количество витков спирали у человека – от 5 до 14.
По морфологии сперматозоиды делятся на три типа:
примитивный тип. Мелкие жгутиковые клетки с округлым или коническим ядром, в области шейки, или средней части, обычно вокруг основания жгутика располагаются 4 митохондрии. Этот тип приспособлен к наружному оплодотворению яиц в воде (губки, многие рыбы и многие виды беспозвоночных животных).
Модифицированный тип (у млекопитающих). Ядро и головка вытянуты, число митохондрий сильно увеличено, и расположены они особым образом в виде оболочки, встречаются ундулирующие мембраны.
Сложный тип (у ракообразных , нематод и других беспозвоночных) – безжгутиковые. Округлой или звездчатой формы, могут перемещаться с помощью псевдоподий или оставаться неподвижными.
Как правило сперматозоиды – очень мелкие клетки. Однако известны и гигантские спермии: у дрозофилы Drosophila bifurca длина сперматозоида составляет 58 мм, он в 20 раз длиннее тела взрослой особи и в 15 000 раз длиннее спермия человека.
В результате мейоза из одного сперматоцита I порядка образуется 4 зрелых сперматиды.
Мейоз проходит без перерывов.
Дифференцировка проходит после мейотических делений в период формирования (дополнительный период по сравнению с оогенезом).
В результате неполной цитотомии клетки развиваются в составе синцитиального клона.
На стадии диплотены транскрибируются в основном гены, продукты которых необходимы для обеспечения подвижности спермия или для связывания его с яйцом (тубулин, биндины), а также протамины. Также показана транскрипция генов в раннем спермиогенезе, за счет этого все спермии имеют разный фенотип. Важными для спермиогенеза являются такие гены, как crem, adam (у млекопитающих) и don juan (у дрозофилы).
Сперматогенез можно разделить на два типа: алиментарный, происходящий с участием вспомогательных соматических клеток, и солитарный, присутствующий у животных, сперматозоиды которых развиваются без связи с соматическими клетками.
Различные типы строения семенника
Первым рассмотрим канальцево-цистный тип строения (у костистых рыб, рис. 3.). Так же, как и в случае канальцевого типа, здесь зачатковая зона семенника (стволовые сперматогонии) находятся внутри канальцев. Однако в канальцево-цистных семенниках степень интеграции сперматогенного эпителия меньше, чем в канальцевых семенниках Amniota: фолликулярный эпителий гетерогенен по своему морфофизиологическому состоянию в зависимости от стадии развития связанных с ним половых клеток (клетки цист, сперматофоры, свободные фолликулярные клетки).
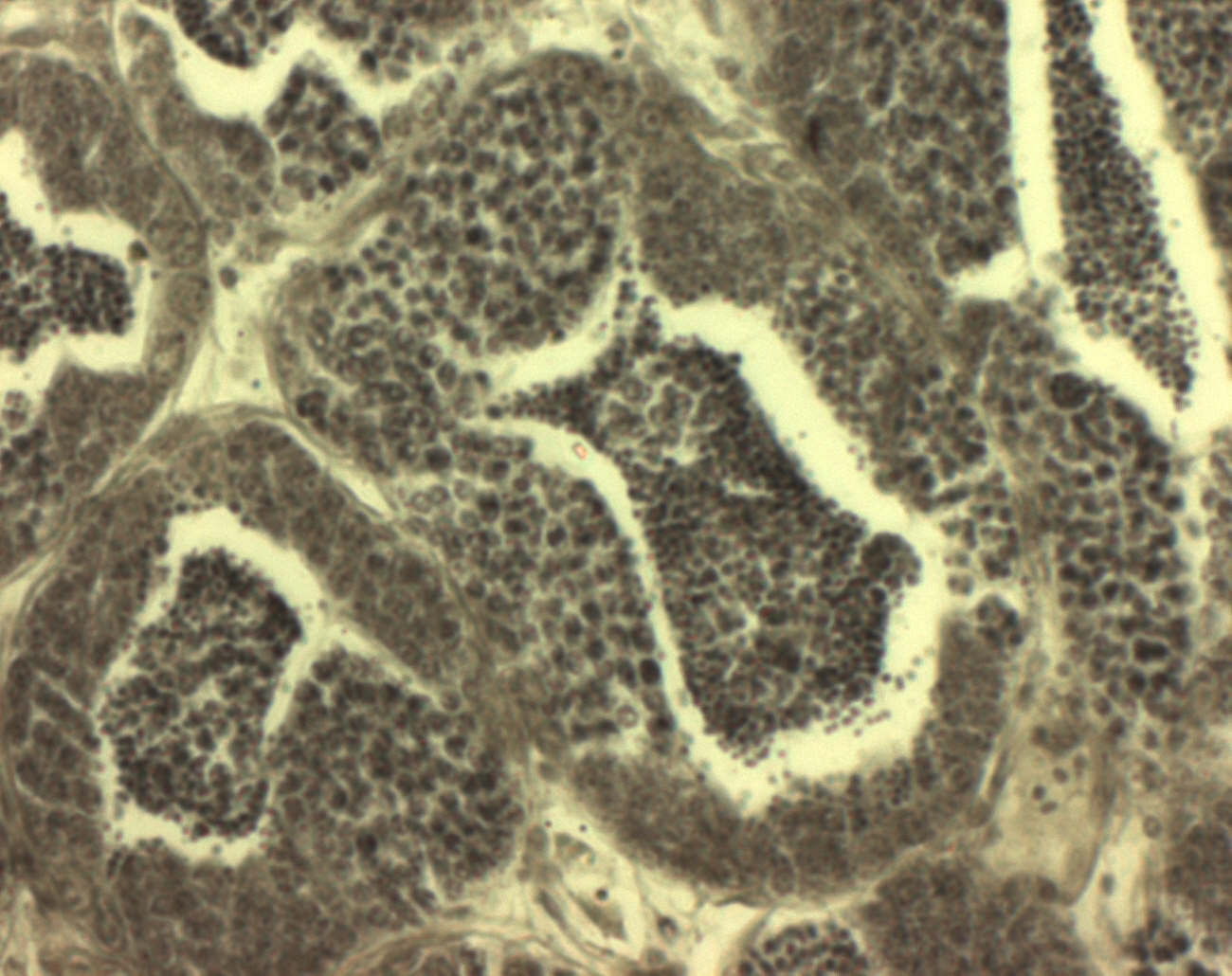
Рис.3 Срез семенника псевдорасбора (увеличение х40).
Далее мы рассмотрим фолликулярно-цистный тип семенника (на примере хвостатых амфибий). У хрящевых рыб стенка цисты образована одной клеткой, позже в филогенезе, очевидно, возникли многоклеточные цисты, требующие межклеточной интеграции в функционировании фолликулярных клеток. Канальцы в данном типе семенников не образуются, вместо них – отдельные фолликулы, внутри которых расположены цисты (рис. 4.).

Рис.4 Срез семенника тритона (увеличение х20).
Самым древним типом организации семенника считается фолликулярный тип, который характеризуется наиболее примитивными взаимоотношениями половых и вспомогательных клеток, и описан у круглоротых. В этом случае отростки клеток фолликулярного эпителия, выстилающего полость фолликулов, разветвляются между половыми клетками, не образуя упорядоченных клеточных ассоциаций, а развитие сперматид осуществляется без контакта с фолликулярным эпителием.
В случае фолликулярного и фолликулярно-цистного типов строения семенника стволовые сперматогонии расположены за пределами фолликулов, в соединительнотканной строме семенника. Объединение всех клеток полового зачатка в единой системе сперматогенного эпителия у высших Anamnia и Amniota создало более стабильные условия для их развития.
Cперматогенез у млекопитающих
Сперматогенез – это процесс постепенной трансформации сперматогониальных стволовых клеток в сперматозоиды в течение длительного периода времени в пределах границ семеносных канальцев семенника .
Первичные половые клетки позвоночных, оказавшись в гонадных валиках мужского зародыша, включаются в состав половых тяжей. Здесь они остаются до созревания; к этому времени в половых тяжах образуются полости, и тяжи превращаются в семенные канальцы, эпителий которых дифференцируется в клетки Сертоли. Инициация сперматогенеза в период полового созревания вероятно регулируется синтезом BMP8B в сперматогенных половых клетках - сперматогониях. Дифференцировка половых клеток начинается после достижения критической концентрации BMP8B. Продукция BMP8B в дифференцирующихся клетках достигает высокого уровня, и тем самым стимулируется дальнейшая дифференцировка этих клеток. У половозрелых мышей, лишенных BMP8B, сперматогенез не начинается.
Ст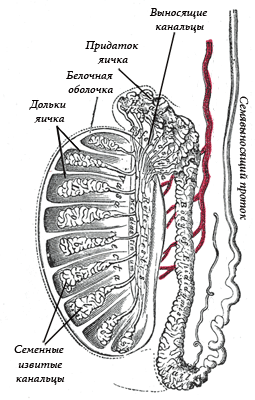 роение
семенника
роение
семенника
Рис. 1. Строение семенника млекопитающих.
Снаружи большая часть семенника покрыта серозной оболочкой (брюшиной), под которой расположена плотная соединительнотканная оболочка – белочная оболочка (tunica albuginea). На заднем крае семенника белочная оболочка утолщается, формируя средостение, от которого вглубь семенника отходят соединительнотканные перегородки, разделяющие его на дольки (Рис. 1). Каждая долька содержит семенные канальцы, промежутки между которыми заполнены соединительной тканью. Семенной каналец состоит из длинной извитой части, где происходит сперматогенез, и короткой прямой части, ведущей к средостению и соединяющийся там с соединяющимися друг с другом канальцами сети семенника. Стенки извитой части семенных канальцев содержат расположенные в несколько рядов мужские половые клетки на разных стадиях сперматогенеза, а также клетки Сертоли, обеспечивающие питание половых клеток. По мере развития половые клетки продвигаются от наружной поверхности семенных канальцев к их просвету. В промежутках между семенными канальцами среди нежной соединительной ткани долек расположены группы интерстициальных клеток Лейдига, выделяющих мужской половой гормон - тестостерон.
Придаток семенника является частью выводного протока. Он служит для накапливания зрелых спермиев, которые могут сохраняться в нем, в течение довольно длительного времени в неподвижном состоянии. Секрет придатков обеспечивает сохранение жизнеспособности и подвижности спермиев. В процессе спаривания животных перистальтические сокращения мышц придатка выбрасывают спермии в семяпровод.
Рассмотрим процесс сперматогенеза на примере грызунов (мыши).

Рис. 2. Семенник мыши (Х40). Эозин.
Весь процесс можно разделить на 3 основные стадии: стадия размножения или пролиферации при помощи митотических делений, в которой участвуют сперматогонии; редукционные деления или мейоз, в который вступают сперматоциты и стадия дифференцировки или спермиогенеза, результатом которой является образование сперматозоидов из сперматид.
Первая стадия - фаза размножения (пролиферации), в этот период клетки носят название сперматогоний, которые путем делений и дифференцировки превращаются в первичные сперматоциты.
Для мышей сперматогонии являются сперматогониальными стволовыми клетками (ССК) и носят название сперматогиний типа Аизолированная (Аsingle). Эта клетка лежит изолированно от других сперматогоний, имеет овальное ядро с диффузным деконденсированным хроматином, равномерно распределенным по ядру, с одним или двумя четко очерченными ядрышками. Бедная органеллами цитоплазма стволовых сперматогониальных клеток содержит незначительное число митохондрий, слабо развитый эндо-плазматический ретикулум и аппарат Гольджи. В то же время в цитоплазме этих клеток имеется большое число свободных рибосом и полисом. Такие клетки редко делятся.
Согласно теории Оакберга и Хаккинса [Oakberg, Huckins, 1976] периодические деления сперматогоний типа Аsingle, приводят либо к появлению таких же клеток сперматогоний типа Аsingle, либо к образованию клеток, которые не могут завершить цитокинез и остаются соединенными цитоплазматическими мостиками. Такие группы пар сперматогоний носят название сперматогонии типа Аспаренные (Apaired). В результате деления сперматогоний типа Apaired образуются цепочки, или группы клеток, соединенных цитоплазматическими мостиками, которые называются сперматогониями типа Агрупповые (Аaligned). Эти сперматогонии трансформируются в сперматогонии типа А1. Далее у крыс и мышей происходит шесть последовательных делений, в результате которых сменяют друг друга сперматогонии типа А1 — А4, промежуточного типа (In) и типа B (рис. 3).

Рис. 3. Схема развития мужских половых клеток мышей. ССК – сперматогониальная стволовая клетка; Apr – Apaired; Aal – Aaligned; A1-A4 - сперматогонии типа А; In - промежуточного типа; В – сперматогонии типа B; пЛ – прелоптотема; Л – лептотема; З – зиготема; П – пахитема; Дип – диплотема; Д- диакинез; СПII - сперматоцит II порядка; ОС – округлая сперматида; 5-15 – последовательные генерации в ходе спермиогенеза.
Таким образом, можно выделить три основных клеточных компартмента:
Компартмент обновляющихся клеток, содержащий сперматогоний типа Аsingle, подразделяются на две субпопуляции: сперматогоний типа Аsingle — долгоживущие, и быстрообновляющиеся стволовые клетки, которые делятся один раз в течение цикла сперматогенного эпителия;
Компартмент недифференцированных клеток, включающий сперматогоний типа Apaired и типа Аaligned, которые в отличие от сперматогоний типа Аsingle соединены цитоплазматическими мостиками;
Компартмент, включающий дифференцирующиеся клетки, куда относят сперматогоний типа A1 — А4, In и типа B.
Сперматогогии типа В являются последней генерацией, которые претерпевают митотические деления, при этом формируя две клетки следующей стадии – сперматоциты, которые находятся на стадии прелептотены и мигрируют в сторону просвета канальца через плотные контакты клеток Сертоли. С этой стадии можно говорить о начале мейотического деления.
В период предмейотической интерфазы происходит синтез ДНК, и клетки в начале мейоза содержат удвоенное количество ДНК (4с).
Профаза мейоза делится на следующие стадии: прелептотему, лептотему, зиготему, пахитему, диплотему и диакинез. В прелептотему происходит удвоение ДНК. На стадии лептотемы в ядре появляются хромосомы в виде тонких нитей, число их равно диплоидному количеству. На стадии зиготемы начинается попарная конъюгация гомологичных хромосом с формированием синаптонемных комплексов (СК), которые являются частью бивалента. Стадия пахитемы — это стадия толстых нитей, конъюгирующие хромосомы становятся спирализованными. Объединение двух конъюгирующих хромосом называется бивалентом. На этой стадии происходит кроссинговер – обмен участками между гомологичными хромосомами. На стадии диплотемы тесно соединенные хромосомы начинают отталкиваться друг от друга и расходятся. Однако в точках, где прошел кроссинговер, остаются связи — хиазмы. На стадии диакинеза исчезает ядрышко, спирализация хромосом усиливается. В прометафазе I происходит разрушение ядерной оболочки и достигается максимальная спирализация хромосом. В метафазе I конъюгирующие хромосомы располагаются у экватора веретена, и на стадии анафазы I гомологичные хромосомы расходятся к полюсам.
В дочерних клетках — сперматоцитах II — число хромосом уменьшено вдвое (гаплоидное), но масса ДНК соответствует диплоидному набору (2с). За телофазой I идет короткая интерфаза, после которой происходит второе деление созревания. Интерфаза второго деления отличается тем, что в ней не происходит синтез ДНК. В период второго деления созревания сестринские хроматиды каждой хромосомы расходятся к полюсам, и, таким образом, в результате двух мейотических делений из каждого сперматоцита образуются четыре сперматиды с гаплоидным набором хромосом и гаплоидной массой ДНК, которые вступают в следующую стадию – дифференцировки.
Гаплоидные сперматиды — мелкие недифференцированные клетки, которые более не делятся и проходят длительную фазу терминальной дифференцировки, известной как спермиогенез, превращаясь в дифференцированные сперматозоиды. Клетки претерпевают кардинальные изменения, в том числе следующие основные модификации:
Сильное уменьшение объема ядра, его удлинение, конденсация хроматина
Аппарат Гольджи производит лизосомальные гранулы, которые в виде шапочки перемещаются в пространство перед ядром, пока в конце концов не формируют акросому. Акросомальная система содержит гидролитические ферменты, которые необходимы для взаимодействия сперматозоида с яйцеклеткой и оплодотворения.
Клетка образует жгутик с митохондриями в проксимальной области, в формировании жгутика участвует дистальная центроль.
Сперматозоид теряет избыток цитоплазмы почти со всеми органеллами, которые в виде остаточных телец сползают со сперматозоида и фагоцитируются клетками Сертоли.

Рис. 4. Семенник крысы (Х20). Гематоксилин+эозин.
Рис. 5. Семенник крысы (Х40). Гематоксилин.
У позвоночных животных сперматогенные клетки развиваются в карманах, образованных клетками Сертоли — эпителиальными клетками, выстилающими просвет извитых канальцев, обеспечивающими питание половых клеток и. К концу созревания происходит процесс спермиации, или выход сперматозоидов из карманов клеток Сертоли. Дальнейшая дифференциация сперматид (удаление избыточной цитоплазмы, приобретение подвижности) заканчивается в придатке семенника.
Во время сперматогенеза происходит апоптоз, при этом значительная часть клеток дегенерирует в процессе сперматогониальной стадии, называемой «плотность – зависимая регуляция», преимущественно при митотических делениях сперматогоний типа А2 и А4. Одна из возможных причин, что дегенерация это гомеостатический механизм для ограничения числа клеток, которые могут поддерживаться с помощью клеток Сертоли. Апоптоз также часто происходит во время мейоза, что, возможно, связано с повреждениями хромосом.
Стенку канальца составляет т.н. сперматогенный эпителий, сформированный разными по происхождению клеками: соматическими клетками гонады и клетками полового зачатка (Рис. 6). Роль соматических клеток играют клетки Сертоли, которые поддерживают и питают созревающие половые клетки на всех стадиях их развития, формируют стенку канальца, а также образуют нишу для ССК. Основания клеток Сертоли, прилежащие к базальной мембране семенных канальцев, разделены группами сперматогоний и прелептотенных сперматоцитов. Таким образом, базальная область фолликулярного эпителия представляет собой аркады клеток Сертоли, сомкнутые над прилегающими к базальной мембране группами половых клеток1. В участках контактов клеток Сертоли имеют место межклеточные соединения типа плотных контактов (tight junctions).
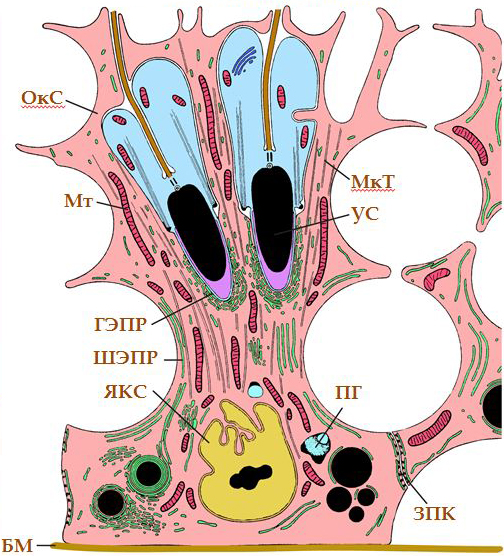
Рис. 6. Просвет семенного канальца. БМ – базальная мембрана; ГЭПР – гладкий эндоплазматический ретикулум; МкТ – микротрубочка; Мт – митохондрия; ПГ – пигментная гранула; РЦ – ризидуальная цитоплазма сперматид; УС – удлиняющаяся сперматида; ШЭПР – шероховатый эндоплазматический ретикулум; ЯКС – ядро клетки Сертоли.
Зоны плотных контактов между клетками Сертоли обусловливают разделение сперматогенного эпителия на два отсека (компартмента): базальный (в нем между основаниями сустентоцитов расположены сперматогонии и прелептотенные сперматоциты) и адлюменальный, находящийся выше уровня этих зон, в котором заключены все более поздние генерации половых клеток. В процессе сперматогенеза периодически происходит перемещение сперматоцитов из базального отсека в адлюменальный — через зоны плотных контактов. Эта транслокация осуществляется без нарушения замкнутости барьера между отсеками. Отростки фолликулярных клеток, внедряясь в базальный отсек, отделяют очередную группу сперматоцитов от базальной мембраны. В области соприкосновения отростков друг с другом происходит новообразование специализированного плотного контакта, площадь которого, постепенно увеличиваясь в высоту, смещает расположенную над ним группу половых клеток в направлении полости канальцев. Размыкание межклеточного контакта над перемещающейся группой сперматоцитов завершает их транслокацию в адлюменальный отсек. Полагают, что половые клетки пассивны в этом перемещении, а описанные изменения межклеточных контактов служат механизмом их передвижения из одного компартмента сперматогенного эпителия в другой.
Половые клетки развиваются в синцитии, соединенные через цитоплазматические мостики. Этот уникальный процесс неполного деления обеспечивает синхронное развитие и позволяет осуществляться быстрой связи между развивающимися клетками.
Клетки Сертоли играют важную роль в сперматогенезе:
Поддежание и питание половых клеток
Компартментализация семенного канальца с помощью плотных контактов (основной компонент гематотестикулярного барьера, который препятствует проникновению в адлюменальный отсек сперматогенного эпителия и в полость семенных канальцев белков, аминокислот, гонадотропинов, иммуноглобулинов, осмотически активных веществ, витальных красителей и т. д. Этот барьер, проходящий на уровне специализированных контактов фолликулярного эпителия, создает в полости семенных канальцев и прилежащем к ней отсеке сперматогенного эпителия особую среду, в значительной степени изолированную от гуморальных влияний организма, и благоприятную для осуществления мейоза и завершения развития сперматозоидов)
Контролируемое высвобождение зрелых сперматид в просвет канальца (спермиация)
Фагоцитоз дегенерирующих половых клеток и излишней цитоплазмы (остаточных телец), которая сползает со зрелого сперматозоида
Секреция белков, факторов роста
Хорошо известна тесная зависимость эндокринных функций семенника от воздействия гонадотропных гормонов гипофиза. При воздействии ФСГ, клетки Сертоли синтезируют андрогенсвязывающий белок, ответственный за поддержание в них определенного уровня содержания андрогенов. С другой стороны, клетки фолликулярного эпителия семенников вырабатывают ингибин, тормозящий продукцию аденогипофизом ФСГ, что свидетельствует о существовании обратной связи в регуляции стероидогенеза между фолликулярным эпителием и гипофизом.