Генотип как целостная система
Свойства генов. На основании знакомства с примерами наследования признаков при моно- и дигибридном скрещивании может сложиться впечатление, что генотип организма слагается из суммы отдельных, независимо действующих генов, каждый из которых определяет развитие только своего признака или свойства. Такое представление о прямой и однозначной связи гена с признаком чаще всего не соответствует действительности. На самом деле существует огромное количество признаков и свойств живых организмов, которые определяются двумя и более парами генов, и наоборот, один ген часто контролирует многие признаки. Кроме того, действие гена может быть изменено соседством других генов и условиями внешней среды. Таким образом, в онтогенезе действуют не отдельные гены, а весь генотип как целостная система со сложными связями и взаимодействиями между ее компонентами. Эта система динамична: появление в результате мутаций новых аллелей или генов, формирование новых хромосом и даже новых геномов приводит к заметному изменению генотипа во времени.
Характер проявления действия гена в составе генотипа как системы может изменяться в различных ситуациях и под влиянием различных факторов. В этом можно легко убедится, если рассмотреть свойства генов и особенности их проявления в признаках:
Ген дискретен в своем действии, т. е. обособлен в своей активности от других генов.
Ген специфичен в своем проявлении, т. е. отвечает за строго определенный признак или свойство организма.
Ген может действовать градуально, т. е. усиливать степень проявления признака при увеличении числа доминантных аллелей (дозы гена).
Один ген может влиять на развитие разных признаков — это множественное, или плейотропное, действие гена.
Разные гены могут оказывать одинаковое действие на развитие одного и того же признака (часто количественных признаков) — это множественные гены, или полигены.
Ген может взаимодействовать с другими генами, что приводит к появлению новых признаков. Такое взаимодействие осуществляется опосредованно — через синтезированные под их контролем продукты своих реакций.
Действие гена может быть модифицировано изменением его местоположения в хромосоме (эффект положения) или воздействием различных факторов внешней среды.
Взаимодействия аллельных генов. Явление, когда за один признак отвечает несколько генов (аллелей), называется взаимодействием генов. Если это аллели одного и того же гена, то такие взаимодействия называются аллельными, а в случае аллелей разных генов —неаллельными.
Выделяют следующие основные типы аллельных взаимодействий: доминирование, неполное доминирование, сверхдоминирование и кодоминирование.
Доминирование —тип взаимодействия двух аллелей одного гена, когда один из них полностью исключает проявление действия другого. Такое явление возможно при следующих условиях: 1) доминантный аллель в гетерозиготном состоянии обеспечивает синтез продуктов, достаточный для проявления признака такого же качества, как и в состоянии доминантной гомозиготы у родительской формы; 2) рецессивный аллель совсем неактивен, либо продукты его активности не взаимодействуют с продуктами активности доминантного аллеля.
Примерами такого взаимодействия аллельных генов может служить доминирование пурпурной окраски цветков гороха над белой, гладкой формы семян над морщинистой, темных волос над светлыми, карих глаз над голубыми у человека и т. д.
Неполное доминирование, или промежуточный характер наследования, наблюдается в том случае, когда фенотип гибрида (гетерозиготы) отличается от фенотипа обеих родительских гомозигот, т. е. выражение признака оказывается промежуточным, с большим или меньшим уклонением в сторону одного или другого родителя. Механизм этого явления состоит в том, что рецессивный аллель неактивен, а степень активности доминантного аллеля недостаточна для того, чтобы обеспечить нужный уровень проявления доминантного признака.
Примером неполного доминирования является наследование окраски цветков у растений ночной красавицы (рис. 3.5). Как видно из схемы, гомозиготные растения имеют либо красные (АА), либо белые (аа) цветки, а гетерозиготные (Аа) — розовые. При скрещивании растения с красными цветками и растения с белыми цветками в F1, у всех растений цветки розовые, т. е. наблюдается промежуточный характер наследования. При скрещивании гибридов с розовой окраской цветков в F2 имеет место совпадение расщепления по фенотипу и генотипу, так как доминантная гомозигота (АА) отличается от гетерозиготы (Аа). Так, в рассматриваемом примере с растениями ночной красавицы расщепление в F2 по окраске цветков обычно следующее — 1 красная(АА): 2 розовые (Аа): 1 белая (аа).
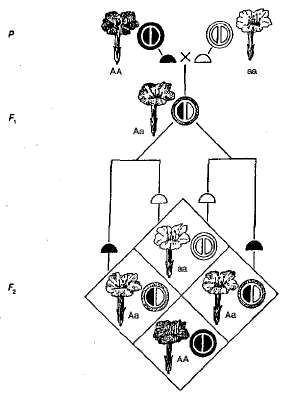
Рис. 3.5. Наследование окраски цветков при неполном доминировании у ночной красавицы.
Неполное доминирование оказалось широко распространенным явлением. Оно наблюдается в наследовании курчавости волос у человека, масти крупного рогатого скота, окраски оперения у кур, многих других морфологических и физиологических признаков у растений, животных и человека.
Сверхдоминирование — более сильное проявление признака у гетерозиготной особи (Аа), чем у любой из гомозигот (АА и аа).Предполагается, что это явление лежит в основе гетерозиса (см. § 3.7).
Кодаминирвание— участие обоих аллелей в определении признака у гетерозиготной особи. Ярким и хорошо изученным примером кодоминирования может служить наследование IV группы крови у человека (группа АВ).
Эритроциты людей этой группы имеют два типа антигенов: антиген А (детерминируемый геном /\ имеющимся в одной из хромосом) и антиген В (детерминируемый геном /а, локализованным в другой гомологичной хромосоме). Только в этом случае проявляют свое действие оба аллеля — 1А (в гомозиготном состоянии контролирует II группу крови, группу А) и IB (в гомозиготном состоянии контролирует III группу крови, группу В). Аллели 1А и IB работают в гетерозиготе как бы независимо друг от друга.
Пример наследования групп крови иллюстрирует и прояв-ление множественного аллелизма: ген/может быть представлен тремя разными аллелями, а есть гены, имеющие десятки аллелей. Все аллели одного гена получили название серии мно-жественных аллелей, из которых каждый диплоидный организм может иметь два любых аллеля (и только). Между этими аллелями возможны все перечисленные варианты аллельных взаимодействий.
Явление множественного аллелизма распространено в природе. Известны обширные серии множественных аллелей, определяющих тип совместимости при оплодотворении у грибов, опылении у семенных растений, детерминирующих окраску шерсти животных и т. д.
Взаимодействия неаллельных генов.Неаллельные взаимодействия генов описаны у многих растений и животных. Они приводят к появлению в потомстве дигетерозиготы необычного расщепления по фенотипу: 9:3:4; 9:6:1; 13:3; 12:3:1; 15:1, т.е. модификации общей менделевской формулы 9:3:3:1. Известны случаи взаимодействия двух, трех и большего числа неаллельных генов. Среди них можно выделить следующие основные типы: комплементарность, эпистаз и полимерию.
Комплементарным, или дополнительным, называется такое взаимодействие неаллельных доминантных генов, в результате которого появляется признак, отсутствующий у обоих родителей. Например, при скрещивании двух сортов душистого горошка с белыми цветками появляется потомство с пурпурными цветками. Если обозначить генотип одного сорта ААbb, а другого — ааВВ,то
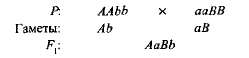
Гибрид первого поколения с двумя доминантными генами (А и В) получил биохимическую основу для выработки пурпурного пигмента антоциана, вто время как поодиночке ни ген А, ни ген B не обеспечивали синтез этого пигмента. Синтез антоциана представляет собой сложную цепь последовательных биохимических реакций, контролируемых несколькими неаллельными генами, и только при наличии как минимум двух доминантных генов (А-В-) развивается пурпурная окраска. В остальных случаях{ааВ- и A-bb) цветки у растения белые (знак «—» в формуле генотипа обозначает, что это место может занять как доминантный, так и рецессивный аллель).
При самоопылении растений душистого горошка из F1 в F2 наблюдалось расщепление на пурпурно- и белоцветковые формы в соотношении, близком к 9:7. Пурпурные цветки были обнаружены у 9/16 растений, белые — у 7/16. Решетка Пеннета наглядно показывает причину этого явления (рис. 3.6).
Эпистаз — это такой тип взаимодействия генов, при котором аллели одного гена подавляют проявление аллельной пары другого гена. Гены, подавляющие действие других генов, называются эпистатическими, ингибиторами или супрессорами. Подавляемый ген носит название гипостатический.
По изменению числа и соотношения фенотип и чес ких классов при дигибридном расщеплении в F2 рассматривают несколько типов эпистатических взаимодействий: доминантный эпистаз (А>В или В>А) с расщеплением 12:3:1; рецессивный эпистаз (а>Вили b>А), который выражается в расщеплении 9:3:4, и т. д.
Полимерия проявляется в том, что один признак формируется под влиянием нескольких генов с одинаковым фенотипичес-ким выражением. Такие гены называются полимерными. В этом случае принят принцип однозначного действия генов на развитие признака. Например, при скрещивании растений пастушьей сумки с треугольными и овальными плодами (стручочками) в F1образуются растения с плодами треугольной формы. При их самоопылении в F2 наблюдается расщепление на растения с треугольными и овальными стручочками в соотношении 15:1. Это объясняется тем, что существуют два гена, действующих однозначно. В этих случаях их обозначают одинаково— А1и A2 .
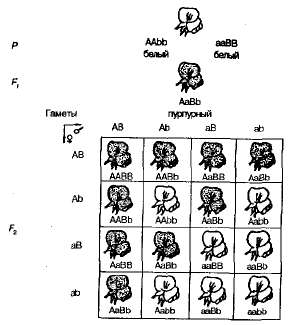
Рис. 3.6. Наследование окраски цветков у душистого горошка
Тогда все генотипы (А1 ,-А2,-, А1-а2а2, a1a1A2-) будут иметь одинаковый фенотип — треугольные стручочки, и только растенияа1а1а2a2 будут отличаться —- образовывать овальные стручочки. Это случай некумулятивной полимерии.
Полимерные гены могут действовать и по типу кумулятивной полимерии. Чем больше подобных генов в генотипе организма, тем сильнее проявление данного признака, т. е. с увеличением дозы гена (А1 А2 А3 и т. д.) его действие суммируется, или кумулируется. Например, интенсивность окраски эндосперма зерен пшеницы пропорциональна числу доминантных аллелей разных генов в тригибридном скрещивании. Наиболее окрашенными были зерна А1А1А2А2А3,А 3 а зерна а1а1а2a2а3а 3 не имели пигмента.
По типу кумулятивной полимерии наследуются многие признаки: молочность, яйценоскость, масса и другие признаки сельскохозяйственных животных; многие важные параметры физической силы, здоровья и умственных способностей человека; длина колоса у злаков; содержание сахара в корнеплодах сахарной свеклы или липидов в семенах подсолнечника и т. д.
Таким образом, многочисленные наблюдения свидетельствуют о том, что проявление большей части признаков представляет собой результат влияния комплекса взаимодействующих генов и условий внешней среды на формирование каждого конкретного признака.

Роль генотипа и условий внешней среды в формировании фенотипа
Все многообразие всего живого и его постоянное совершенствование были бы невозможны без изменчивости. Это связано с тем, что генотип последовательно реализуется в фенотип в ходе индивидуального развития организма и в определенных условиях среды обитания, факторы которой (колебания освещенности, температуры, влажности, условий питания, взаимоотношений с другими организмами и др.) часто оказывают определяющее значение на проявление и развитие того или иного признака и свойства. Поэтому организмы, имеющие одинаковые генотипы, могут заметно отличаться друг от друга по фенотипу. Приведем несколько примеров.
Если растения примулы, которые в обычных условиях имели красные цветки, перенести в оранжерею и содержать их там при температуре 30—35°С и высокой влажности, то через некоторое время все цветки у них оказываются белыми. Если эти же растения вновь поместить в обычные (комнатные) условия, то распустившиеся цветки будут красными.
Другим классическим примером, иллюстрирующим влияние внешней среды на проявление качественных признаков, служит изменение окраски шерсти у гималайского кролика. Обычно при 20°С у этого кролика шерсть белая, за исключением черных ушей, лап, хвоста и мордочки. При 30°С такие кролики вырастают полностью белыми. Если же у гималайского кролика сбрить шерсть на боку или спине и содержать его при температуре воздуха ниже +2°С, то вместо белой шерсти вырастет черная. Но если сбрить шерсть на ухе, то в обычных условиях там снова вырастет черная шерсть.
Эти наблюдения объясняют, почему гималайские кролики рождаются белыми, без участков черной шерсти: их эмбриональное развитие происходит в условиях высокой температуры.
Известно, что все признаки и свойства организма наследственно детерминированы, однако организмы наследуют не сами признаки и свойства, а лишь возможность их развития. Для проявления и развития признака необходимы соответствующие условия внешней среды.
Но даже в том случае, когда проявление и развитие того или иного признака происходит, степень его выраженности бывает
разной в зависимости от условий внешней среды: при одних она усиливается, при других ослабляется. Пределы этих изменений определены возможностями, заложенными в генотипе.
Пределы модификационной изменчивости признака, ограниченные действием генотипа, называют его норной реакции.
Для разных признаков и свойств организмов границы, определяемые нормой реакции, неодинаковы. Наибольшей пластичностью и изменчивостью характеризуются количественные признаки: семенная продуктивность злаков, величина удоя у крупного рогатого скота, масса животных, число и размеры листьев и колосков у растений и т. д. В то же время качественные признаки (масть животных, окраска семян, цветков и плодов, остистость и опушенность колоса у растений) мало зависят от условий среды.
Так, у крупного рогатого скота удой во многом определяется условиями его содержания. При подборе кормов нужного качества и количества ценная порода скота может дать 5—6 тыс. кг молока в год, а в случае ухудшения условий содержания продуктивность снижается до 2,5 тыс. кг и даже ниже. Что касается масти животных, то при самых разных условиях она почти не изменяется.
Подобную закономерность можно наблюдать и у растений. Например, растения элитного сорта картофеля, выращенные в условиях высокой культуры агротехники, дают урожай клубней 500—600 ц/га и более. Ухудшение условий (тяжелые глинистые почвы, низкая культура земледелия и ухода за растениями) может привести к тому, что урожайность прекрасного сорта картофеля снизится до 100—150 ц/га и ниже. Правда, окраска клубней и их форма при этом изменяются в очень малой степени.
Рассмотренные примеры свидетельствуют о том, что количественные признаки обладают очень широкой нормой реакции, а качественные — более узкой.
Таким образом, все признаки и свойства каждого сорта растений и каждой породы животных могут изменяться только в пределах нормы реакции. Поэтому попытки добиться высоких урожаев, резко повысить продуктивность сельскохозяйственных животных только за счет повышения уровня культуры земледелия, улучшения условий кормления и содержания животных несостоятельны. В первую очередь необходимо получить новые сорта растений и породы животных улучшенного генотипа, отзывчивые на хорошие условия возделывания и выращивания.
По механизму возникновения и характеру изменений признаков различают два типа изменчивости — наследственную и ненаследственную.
Под наследственной изменчивостью понимают способность к изменениям самого генетического материала, а под ненаследственной (модификационной, или фенотипической) — способность организмов реагировать на условия окружающей среды и изменяться в пределах нормы реакции, определяемой генотипом.

Модификационная изменчивость
Модификации. Модификациями называют изменения фенотипа, вызванные влиянием окружающей среды и не связанные с изменениями генотипа. При этом возникшее конкретное модифицированное изменение признака не наследуется, но диапазон такой изменчивости, или норма реакции, генетически детерминирована и наследуется. Модификации сохраняются лишь на протяжении жизни данного организма.
Модификационной изменчивости подвержены как количественные, так и качественные признаки. Возникновение модификаций связано с тем, что такие важнейшие факторы среды, как свет, тепло, влага, химический состав и структура почвы, воздух, воздействуют на активность ферментов и в известной мере изменяют ход биохимических реакций, протекающих в развивающемся организме. Этим, в частности, объясняется появление различной окраски цветков у примулы и шерсти у гималайских кроликов, о чем говорилось выше.
Примерами модификационной изменчивости у человека могут служить усиление пигментации кожи (загар) под влиянием ультрафиолетовых лучей, мощное развитие костно-мышечной системы в результате физических нагрузок и т. д. К модификационной изменчивости следует отнести также и явление физиологического гомеостаза — способности организмов противостоять колеблющимся условиям среды путем приспособительного реагирования. Так, у человека при пребывании на разных высотах над уровнем моря вырабатывается неодинаковое количество эритроцитов: в 1 mmj крови у людей, живущих в местностях на уровне моря, их в два раза меньше, чем у людей, живущих высоко в горах.
Число эритроцитов растет пропорционально подъему над уровнем моря. Это явление можно легко объяснить, если вспомнить, что главная функция эритроцитов — перенос кислорода от легких к тканям и углекислого газа от тканей к легким. Увеличение высоты над уровнем моря сопровождается снижением концентрации кислорода в атмосфере, что приводит к недостатку его в тканях. Поэтому насущная потребность в кислороде заставляет человека и животных адаптивно реагировать путем изменения числа эритроцитов на разных высотах.
Эта реакция обратима: переезд в места, расположенные на уровне моря, приводит к снижению числа эритроцитов в крови.
Статистический анализ модификационной изменчивости. Такие условна среды, как влажность, температура, освещенность, физические свойства почвы и ее плодородие, глубина заделки семян, взаимодействие и конкуренция растений с другими сообита-телями, никогда не бывают тождественными даже на одном поле. Поэтому длина колосьев пшеницы на одном поле может колебаться от 6 до 14 см, а размеры листьев одного дерева иногда варьируют в еще более широких пределах, хотя генотип их одинаков. Если листья или колосья расположить в порядке нарастания или убывания их длины, то получается вариационный ряд изменчивости данного признака, слагающийся из отдельных вариант, то есть числа листьев дерева или колосков в колосе пшеницы, имеющих одинаковые показатели.
Как показывают подсчеты, частота встречаемости отдельных вариант в вариационном ряду неодинакова. Чаще всего встречается среднее значение признака, а к обоим концам вариационного ряда частота встречаемости закономерно снижается. Рассмотрим это на примере изменчивости числа колосков в колосе пшеницы. Возьмем произвольно (не выбирая) 100 колосьев одного сорта и подсчитаем в каждом из них число колосков. Полученные цифры (варианты) расположим в порядке нарастания признака и подсчитаем, сколько раз каждая варианта v встречается в каждом ряду р, затем сгруппируем их, т. е. составим вариационный ряд:
![]()
Распределение вариант в этом ряду можно выразить наглядно на графике (рис. 3.12). Для этого на оси абсцисс откладывают значения вариант v в порядке их увеличения, на оси ординат — частоту встречаемости р каждой варианты.
Графическое выражение изменчивости признака, отражающее как размах вариаций, так и частоты встречаемости отдельных вариант, называют вариационной кривой. Установлено, что мо-дификационная изменчивость у растений, животных и человека имеет общие черты.
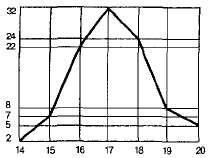
Рис. 3.12. Вариационная кривая числа колосков в колосе пшеницы.
Кривая на графике, как правило, бывает симметричной, особенно когда изучается большое число особей. Это значит, что вариации, как большие, так и меньшие, отличающиеся от среднего арифметического на одну и ту же величину, встречаются одинаково часто. Отсюда следует, что минимальные и максимальные величины должны встречаться очень редко, но с одинаковой частотой.
Значение модификаций.Модификационная изменчивость в естественных условиях носит приспособительный характер и в этом смысле имеет важное значение в эволюции. Обусловленные нормой реакции адаптивные модификации дают возможность организму выжить и оставить потомство в изменившихся условиях среды.
Знание закономерностей модификационной изменчивости имеет также большое практическое значение, так как позволяет предвидеть и заранее планировать максимальное использование возможностей каждого сорта растений и породы животных. В частности, создание заведомо известных оптимальных условий для реализации генотипа обеспечивает их высокую продуктивность.
Такой подход в равной мере относится и к человеку. Каждый ребенок обладает определенными способностями, иногда даже в нескольких областях. Задача психологов и педагогов состоит в том, чтобы как можно раньше найти эту область и обеспечить максимальное развитие ребенка в этом направлении (наряду с общим образованием), т. е. в пределах нормы реакции достичь максимального уровня реализации его генотипа.