

Розділ 2. Природа антигенів
2.1 Антигенність та імуногенність
У визначенні антигену як індуктора імунної відповіді сховані дві його основні характеристики: антигенна специфічність (антигенність), обумовлена його структурними особливостями, і імуногенність - здатність ініціювати імунну систему до формування ефекторів, що нейтралізують антигенну чужорідність.
2.2 Дослідження гаптенів
2.2.1 Гаптени і переносчики (молекули-носії)
Деякі речовини (прості хімічні угруповання) - гаптени - не взмозі забезпечити розвиток імунної відповіді, демонструючи тим самим відсутність властивості имуногенності. Однак вони мають цілком конкретну специфічність – здатність вступати в реакції взаємодії з антитілами, існуючими до їх появи. Карл Ландштейнер вивчав реакцію антитіл на гаптени.
Гаптени набувають імуногенності лише після з’єднання їх з високомолекулярним білком-носієм.
2.2.2 Специфічність серологічних реакцій
Дослідження Ландштейнера виявили принаймні два істотних моменти специфічності антигену:
дуже високий рівень його специфічності (у визначенні специфічності часом бере участь лише один радикал - карбоксильна або аміногрупа);
специфічність високомолекулярного антигену представлена окремими ділянками (епітопами) – місцями зв’язування антитіл або антигенрозпізнаючих рецепторів; при цьому, чим більшою була молекулярна вага антигену, тим більше місць зв’язування було виявлено.
Схема постановки дослідів, розроблена Ландштейнером, включала імунізацію кроликів гаптен-білковим комплексом і наступний аналіз антисироваток від імунізованих тварин з тим же самим або іншим гаптеном, але кон’югованим з іншим неспорідненим білком. Подібний прийом дозволяв працювати тільки з антитілами до гаптену і виключав ті антитіла, що утворювалися до епітопів білка, узятого для імунізації У результаті вдалося показати вирішальну роль тонкої конфігурації гаптена у визначенні специфічності.
У такий спосіб завдяки дослідженням з гаптенами були визначені досить тверді межі для прояву антигенної специфічності і було показано, що незначні структурні розходження між ізомерними формами гаптена визначають утворення різних по специфічності антитіл.
2.2.3 Перехресна реактивність
Модифікований токсин, що не є токсичним, але має імунохімічні характеристики зветься токсоїдом. Так,можемо сказати,що токсоїд імунологічно перехресно реагує з токсином: ці молекули мають спільні епітопи.Імунологічні реакції, в яких імунний компонент (клітина або антитіло) реагує з 2 молекулами які мають спільні епітопи, але в інших аспектах не є подібними, називаються перехресною реактивністю.
2.3 Антитіла до білків
2.3.1 Роль просторової конфігурації і послідовності амінокислот в антигенності
Відповідно до передбачення Ландштейнера вже ранні дослідження показали/підтвердили, що структура молекули білка є важливою для його антигенності (рис. 3). Наприклад, у відповідь на білок утворюються антитіла, які проте не будуть реагувати з цим же білком, який попередньо було денатуровано. Так, антитіла до РНК не реагують з нею, після того як її 4 дисульфідні зв’язки були окислені і відповідно форма (просторова) цієї РНК була змінена.
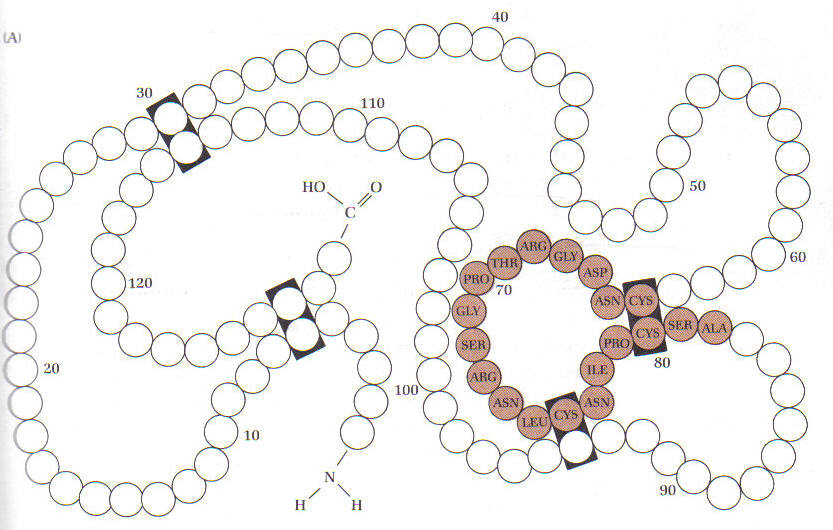
Рисунок 3. Вплив конформації на антигенність