

Ф.Харовіц і матрична теорія
В 1931 році з’явилась нова теорія (рис. 2). Фрідріх Брейн і Фелікс Харовіц імунізували кролів гемоглобіном і проаналізували поєднання амінокислот антитіл, які при цьому продукувались. Вони прийшли до висновку, що відмінність між звичайною молекулою і молекулою антитіла обумовлена розміщенням/порядком аміногруп і формою/просторовою структурою останніх.
Основна провідна думка цієї теорії, що антиген діє як матриця, кожна аміногрупа в антигені буде взаємодіяти,визначати основну групу в антитілі і навпаки.
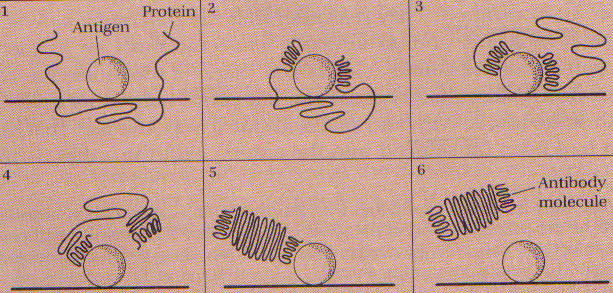
Рисунок 2. Схема Матричної теорії утворення антитіл
Дана теорія порівняно з попередньою була не селективною, а конструктивною,інструктивною. Однак з новими відкриттями – вивченням ДНК, РНК, пептидів і ця теорія була відхилена, оскільки усвідомлення, що амінокислотна послідовність і просторова структура білків є результатом нуклеотидної послідовності генів, які їх кодують, зробили складним/неможливим захист інструктивної моделі утворення антитіл.
1.2.3. Н.Йерне, д.Таллмадж та м.Бернет: Клональна селекція
Тоді Нельс Йерне удосконалив сективну теорію, назвавши її Природною селективною теорією. Дана теорія в основному базувалась на положенні, що всі тварини мають визначений рівень антитіл до широкого спектру антигенів в їх сироватці крові, не зважаючи на те, що вони не були імунізовані даними антигенами. Згідно цієї теорії, антиген проникає в організм і реагує з одним із цих наявних “натуральних” – природніх антитіл в сироватці. Комплекс антиген-антитіло після того транспортується до клітини, і ця клітина є індукованою до продукції саме таких, але багато антитіл. Тобто, знову ж таки – антиген – є селекційний агент.
М.Бернет, який в цей час працював над подібною теорією, запропонував її під назвою Клонально-Селекційна теорія утворення антитіл. Він удосконалив теорію Йерне, запропонувавши такі додаткові уточнення:
Молекули частково денатурованих антитіл можуть стимулювати клітину, здатну продукувати серію її реплікантів (повторів);
Селекція відбувається між антигеном і клітиною, яка продукує антитіла;
Селективна роль антигену полягає в його здатності вибрати відповідний клон лімфоцитів;
Даний лімфоцит має здатність продукувати антитіла однієї або найбільше лише кількох специфічностей;
Лімфоцити експресують антитіла на їх поверхні, які потім реагують з рецептором, отже реакція між антигеном і поверхнею антитіла викликає проліферацію лімфоцитів.
Клон лімфоцитів продукує більше антитіл такої ж специфічності.
1.2.4 Т.Кухн і його замітки про парадигму
Тільки з незначною модифікацією дана теорія змінила статус на парадигму. Парадигма за Т.Кухном відрізняється від теорії тим, що вона не потребує корекції, і є повністю і всіма визнаною.
Сучасний період
Дослідження сучасного періоду дозволяють розуміти/відкрити цілий ряд механізмів у деталях, які лежать в основі імунної відповіді. Це насамперед визначення які клітини її забезпечують, шляхи які лежать в основі їх взаємодії одна з одною, з’ясування природи антитіл, обгрунтування можливості і методів забезпечення такої широкої різноманітності імунної відповіді організму тощо.