

Мембран
В настоящее время с помощью всех новых методов исследования структуры биологических объектов установлено, что огромное значение в создании структуры клеток имеют мембраны (см. рис. 19). Мембраны окружают всю цитоплазму и отграничивают ее от окружающей среды. Проникновение веществ в клетку и из клетки в большой степени зависит от свойств мембраны. Как отметил Д. Бернал, «только после образования мембраны вокруг всей клетки мы действительно имеем то, что с полным правом может быть названо организмом». Кроме того, мембраны образуют оболочки всех органоидов и включений клетки: ядра, митохондрий, лизосом, аппарата Гольджи, эндоплазматического ретикулума.
124
Теории молекулярного строения клеточной мембраны вначале базировались на косвенных данных. Раньше ученые считали, что мембраны состоят только из липидов. Так, Овертон на основании того, что вещества, растворимые в липидах, легко проникают через клеточную мембрану, выдвинул в 1902 г. предположение, что она состоит из тонкого слоя липидов.
Свойства и химический состав мембраны часто изучаются на оболочках эритроцитов. Оболочки эритроцитов представляют собой мембраны, которые очень легко получить путем гемолиза эритроцитов в гипотоническом растворе. В 1925 г. Гортер и Грендел в довольно простых опытах с липидами, экстрагированными из мембран эритроцитов, обнаружили, что площадь монослоя, занимаемого липидами, вдвое больше суммарной площади поверхности всех эритроцитов. Был сделан вывод, что липиды мембран расположены в виде бимолекулярного слоя. Эта теория подкреплялась данными, полученными при измерении электрических параметров клетки, которые свидетельствовали о высоком сопротивлении клеточной мембраны (порядка 1000 Ом/см2). Столь высокое сопротивление характерно для липидов, обладающих малой проводимостью.
Однако многие данные свидетельствовали также о наличии в клеточной мембране белковых молекул. Например, такие свойства мембран, как растяжимость, эластичность и способность некоторых из них к сокращению, можно объяснить лишь наличием в мембране белков, причем фибриллярных белков. Кроме того, при
измерении поверхностного натяжения крупных клеток, например яиц морского ежа, было установлено, что поверхностное натяжение клеток сильно отличается от поверхностного натяжения липидов. Было предположено, что низкое поверхностное натяжение клеточной мембраны обусловлено наличием белковых слоев, покрывающих липидные компоненты мембран. И действительно, добавление к модельной липидно-водной системе небольшого количества белка заметно снижает ее поверхностное натяжение.
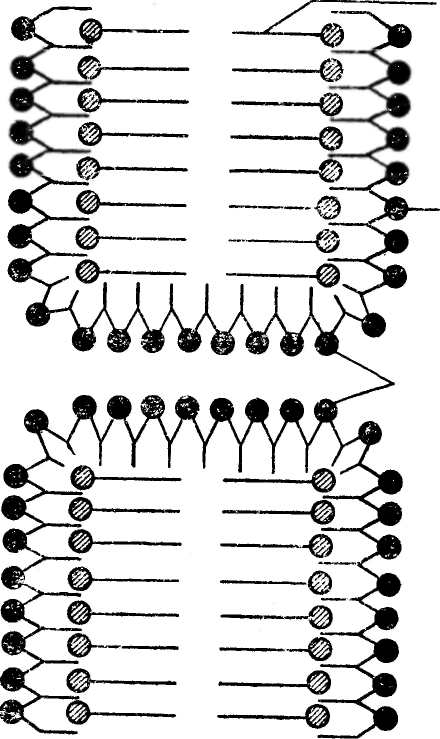
На основе анализа приведенных данных Даниэлли и Давсон в 1935 г. предложили модель строения мембраны, которая не претерпела существенных изменений до нашего времени. Согласно этой модели (рис. 21), имеются два слоя молекул фосфолипидов, которые распо-
125
молекула
липида
молекула
белка
Полярная
пора
Рис.
21. Схема строения клеточной мембраны
по Даниэлли и Давсону.
Гидрофильные концы молекул фосфолипидов содержат полярные группы, которые способны взаимодействовать с дипольными молекулами воды и формировать гидратные оболочки.
На полярных группах молекул фосфолипидов мембраны адсорбированы белковые цепочки, которые в форме глобул покрывают двойной слой фосфолипидов с обеих сторон, придавая ему тем самым известную эластичность и устойчивость к механическим повреждениям, а также низкое поверхностное натяжение. Полярные группы молекул глобулярных белков направлены наружу — в сторону водной фазы, а неполярные группы — в сторону липидов.
К полярным группам относятся аминная, карбоксильная, фосфатная, гидроксильная, карбонильная и некоторые другие.
Поскольку длина липидных молекул равна примерно 3 нм. а толщина монослоя белка не превышает 1 нм, то толщина клеточной мембраны оценивалась примерно в 8 нм.
При этом считалось, что на одну молекулу белка приходится приблизительно 75 — 90 молекул липидов.
Последующие электронномикроскопические исследования подтвердили правильность этой модели. При исследовании ультраструктуры миелина Дж. Робертсоном в
126
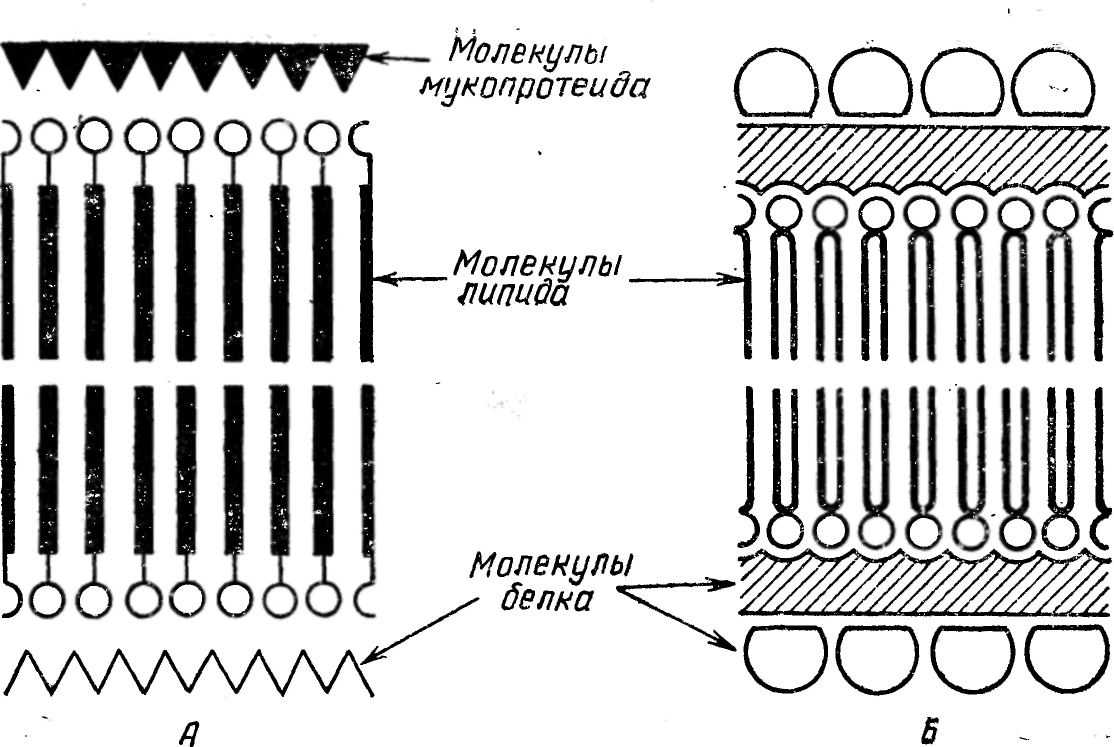
Рис. 22. Схема строения элементарной мембраны.
А — по Дж. Робертсону (1059); Б — современная модель (по Л. Хокину и
М. Хокиной, 1967).
1959 г. была выдвинута теория ординарной или унитарной мембраны. По его мнению, основной единицей всех мембранных структур клеток является трехслойная структура толщиной 7,5 — 9 нм. Эта элементарная мембрана состоит из одного бимолекулярного слоя фосфолипидов, покрытого с цитоплазматическои стороны слоем фибриллярного белка, а с наружной поверхности — мукополисахаридами или мукопротеидами (рис. 22, А).
В настоящее время большинство ученых придерживается следующего представления о структуре мембран: двойной слой фосфолипидов расположен между двумя тонкими слоями молекул фибриллярного белка, которые в свою очередь окружены молекулами глобулярных белков (рис. 22, Б). Белки, входящие в состав мембран, составляют 70 — 75% ее веса (по Гельману) и разделяются на структурные белки, не обладающие ферментативной активностью, и каталитические белки, которые обладают ферментативной активностью.
Способность структурного белка к построению мембран, по мнению Ю. А. Владимирова и Г. И. Клебанова, обусловлена тремя его свойствами: 1) способностью давать комплексы с липидами; 2) способностью стехио-
127
метрически взаимодействовать с другими белками1; 3) способностью к агрегации, которая проявляется в образовании кристаллов, а вместе с фосфолипидами — в участии при образовании мембран.
В настоящее время в литературе широко обсуждается вопрос о природе сил, связывающих мембранные белки и формирующих мембраны. На основе изучения процессов агрегации структурного белка в зависимости от рН среды, ионной силы раствора и других факторов вышеназванные авторы пришли к выводу, что агрегация обусловлена гидрофобными взаимодействиями, скрепляющими субъединицы белка.
В настоящее время большой интерес исследователей вызывает изучение конформации белков мембран. В литературе высказывается предположение, что многие жизненно важные процессы, такие, как окислительное фосфорилирование, активный транспорт веществ, химические реакции при фотосинтезе, проведение нервного импульса, движение цитоплазмы и др., сопровождаются, а может быть, и вызываются изменением способа укладки полипептидной цепи, т. е. изменением конформации белковых молекул в мембранах. Так, в опытах Кеннеди (1967) показано, что активный транспорт некоторых веществ (аминокислот и Сахаров) определяется изменением конформации специфических белков — переносчиков, вмонтированных в мембрану. В связи с этим проводятся очень интенсивные исследования конформации белковых молекул мембран.
При этом применяются методы люминесцентного анализа, инфракрасной спектроскопии, измерения оптической активности и др.
Вторым химическим компонентом мембран являются липиды, составляющие от 20 до 30% их сухого веса. Наибольшая часть липидов представляет собой фосфолипиды, количество которых, например, в митохондриях может достигать 90% от содержания всех липидов. Наиболее распространен в биологических мембранах фосфотидилхолин, но в них содержатся также фосфотидилэтаноламин, фосфотидилинозит и др. Мембраны различных клеток, а также различных органоидов в пределах
1 Стехиометрическое взаимодействие означает такой процесс, при котором имеется вполне определенное количественное соотношение взаимодействующих частиц.
128
одной клетки могут обнаруживать значительное различие в составе липидов.
Раньше считали, что роль фосфолипидов сводятся к приданию мембранам физической структуры и низкой проницаемости для многих веществ. В настоящее время
установлено, что фосфолипиды играют более активную роль. В исследованиях Л. Хокина и М. Хокиной было показано, что если стимулировать прохождение веществ через клеточные оболочки, то этот процесс сопровождается химическими изменениями фосфолипидов — изменением соотношения различных фосфолипидов в мембранах. Обнаруженая метаболическая активность фосфолипидов представляет новое преспективное направлениеисследований.
В последнее время многими учеными развивается глобулярная теория строения мембраны. Во многих случаях на электронных микрофотографиях препаратов мембран, полученных после некоторых видов «нежесткой» фиксации и контрастирования, обнаруживаются глобулярные структуры. На этом основании Шестранд считает, что липидная фаза мембраны существует в виде глобулярных мицелл, представляющих комплекс липидных молекул, залитых в белковый матрикс. Другие ученые считают, что липидные глобулы не полностью окружены белками, а расположены между двумя слоями белка. Все же большинство исследователей (Шестранд Нильсон, Кавана и др.) склоняются к точке зрения, согласно которой субъединицы (блоки) биологических мембран образованы липидными глобулами, полностью окруженными белками и стабилизированным гидрофобным взаимодействием поверхностного слоя белковых молекул. Эти блоки имеют форму правильных шестигранных или пятигранных призм диаметром 8 — 14 нм. Блоки располагаются упорядоченно на расстоянии 8 — 10 нм от центра к центру, образуя в совокупности мембрану. В целом мембрана стабилизирована гидрофобными белок белковыми связями и белок-липидными взаимодействиями. При этом считают, что мембрана не имеет статической организации: форма глобулы в зависимости от функционального состояния мембраны может меняться и становиться или более вытянутой, или более уплощенной. В результате этого мембрана как бы «мерцает» или «пульсирует». В зависимости от той формы, которую принимают глобулы, между ними могут возни-