

Интермедиаты в процессе сворачивания Мультидоменные белки
Обычно уравнения функции распределения используются для расчета модельных функций теплоемкости переходов в тех случаях, когда заселено более двух состояний. Анализ экспериментальных кривых с помощью таких функций является эффективным подходом для детального анализа термодинамических данных дифференциальной сканирующей калориметрии. Точность сегодняшних калориметров такова, что позволяет наблюдать и интерпретировать достаточно малые различия между модельной и экспериментальной кривыми.

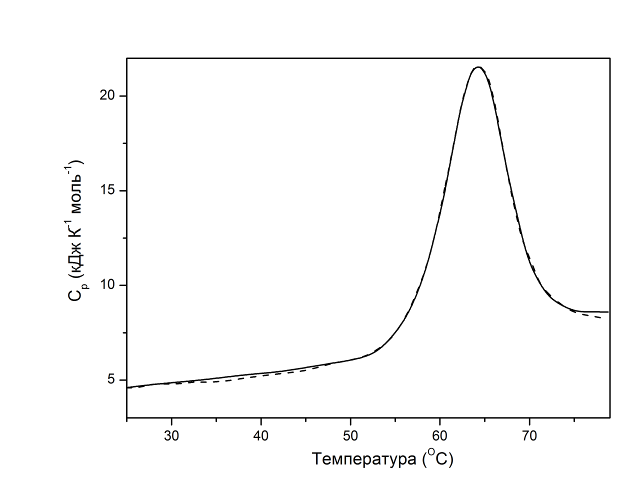
Рис. 15.5. Экспериментальные (сплошные) и модельные (пунктирные) кривые плавления лизоцима куриного яйца при pH 2.5. Пунктирные линии – результат компьютерного моделирования: вверху переход с двумя состояниями, внизу переход с тремя состояниями
Лизоцим куриного яйца является двухдоменным белком и, как свидетельствуют данные, представленные на рисунке 15.5, температурная денатурация обоих его доменов происходит не полностью кооперативно.
Существуют ли заселенные интермедиаты сворачивания в однодоменных белках?
В настоящее время считается, что процесс сворачивания небольших однодоменных белков должен проходить, по крайней мере, через одно переходное состояние, энергия которого выше, чем у нативного и денатурированного состояний. В качестве такого интермедиата была предложена расплавленная глобула, которая, как считают некоторые исследователи, может наблюдаться и в определенных равновесных условиях. Она может образоваться в результате быстрого сворачивания цепи в компактную жидкообразную структуру, которая затем переходит в нативное состояние.
Существование такого состояния позволило бы разрешить парадокс Левинталя, значительно сужая поиск конфигурационного пространства. Суть парадокса состоит в том, что скорость сворачивания полипептидной цепи в уникальную трехмерную белковую структуру, определяемую аминокислотной последовательностью, исключает вероятность того, что этот процесс имеет стохастическую природу, где молекула перебирает огромное число всевозможных конформаций.
Однако свойства расплавленной глобулы достаточно противоречивы. Так, в ней существует развитая вторичная структура, практически в том же количестве, как и в нативном белке. Она слегка менее компактна – её гидродинамический объем всего на 30% больше, чем объем в нативном состоянии. Зато в ней почти полностью отсутствует упорядоченность боковых групп, являющаяся характерной для нативного состояния белка, а довольно высокая скорость водородного обмена в расплавленной глобуле свидетельствует о том, что некоторое количество молекул растворителя могут свободно проникать вглубь неё. Эти противоречия, в известной степени, примиряются предположением о неоднородности расплавленной глобулы, a именно, введением представления о более или менее нативоподобного ядра и опушенной оболочки. Но даже этой простой модели, противоречат некоторые эксперименты по круговому дихроизму и ядерному магнитному резонансу. П. Привалов, анализируя калориметрические данные в поиске интермедиатов процесса сворачивания, сделал вывод, что эти промежуточные состояния представляют собой либо неправильно свернутые формы, образующиеся в неблагоприятных условиях, либо частично развернутые структуры, которые сохранили правильно свернутые участки молекулы. Эти участки обладают всеми характерными чертами кооперативно сворачивающихся доменов. Свободная энергия промежуточнго состояния выше нативного или денатурированного, или их обоих, поэтому его заселенность всегда мала. Главный аргумент против существования расплавленной глобулы полное отсутствие кооперативного плавления.
Сворачивание без барьеров (downhill folding)

а) б)
Рис. 15.6. Схема двустадийной укладки и downhill укладка. а) В двустадийном процессе существует барьер свободной энергии, разделяющий структурированное и неструктурированные состояния б) при downhill укладке барьера свободной энергии не существует
Для подавляющего числа однодоменных белков, изученных до сих пор экспериментально, характерен процесс двустадийного сворачивания, при котором неструктурированное и нативное состояния молекул разделены энергетическим барьером. Однако недавно из теоретических соображений было предположено, что у некоторых белков этот барьер может быть невелик и составлять всего несколько ккал/ моль. Иными словами нативное и неструктурированное состояния белка не разделены сколько-нибудь значительным барьером свободной энергии. Этот барьер может быть еще более понижен за счет либо мутаций, либо изменением состава растворителя. Такое сворачивание получило название без барьерного сворачивания или сворачивания «по склону» (от английского downhill folding). Фактически это означает, что существует такой баланс между энергией и конформационной энтропией, что на профиле поверхностной энергии существует только один минимум для всех отклонений от нативного состояния. Этот минимум может плавно смещается от нативного к развернутому состоянию, например, при изменении температуры (рис. 15.6).
Экспериментальные доказательства существования без барьерного сворачивания белков достаточно скудны и противоречивы, зависят от использованных методов и зачастую одни и те же данные интерпретируются по-разному. Более того, практически для одного и того же белка с незначительными модификациями получены данные как pro так и contra.