

Взаимодействие пептидов и белков с липидами
Липопептиды
Липопептидом принято называть молекулу липида, ковалентно связанную с пептидом. Молекулы известных липопептидов содержат от 4 до 16 аминокислотных остатков. Пептидные цепи при этом могут быть линейными, цикличными (рис. 6.11) или смешанного строения. Иногда в молекуле липопептида могут находиться как L- , так и D-конфигурации одной аминокислоты (рис.6.11).
![]()
Рис.6.11. Структура бацилломицина. D- и L- перед аминокислотами обозначают их различную конфигурацию
Липопротеины (липобелки)
Ряд липидов образуют комплексы со специфическими белками (аполипротеинами); эти комплексы называют липопротеинами. В составе липопротеинов могут быть как свободные жирные кислоты, нейтральные жиры, так и фосфолипиды и холестериды. Молекулы липидов и полипептидов в липопротеинах прочно связаны друг с другом, хотя и не образуют ковалентных связей. Наличие внешней гидрофильной оболочки в липопротеинах делает эти богатые липидами структуры растворимыми в воде и хорошо приспособленными для транспорта липидов из тонкого кишечника в жировые депо и в различные ткани.
Липопротеины значительно различаются по химическому составу, например, соотношение количества белка к количеству липидов в хиломикронах (класс липопротеинов, образующихся в тонком кишечнике в процессе всасывания внешних липидов) составляет примерно 1:99, а у липопротеинов высокой плотности – 50:50. Условно различают свободные или растворимые в водной среде липопротеины (из плазмы крови, молока, желтка яиц и др.), и структурные (мембраны клеток, миелиновой оболочки нервов и др.).
Биомембраны
Клеточные мембраны ограничивают содержимое клетки (или клеточной органеллы) от окружающей среды. Огромная роль мембран в жизненных процессах связана с их относительно большой совокупной площадью. Так, общая площадь всех биологических мембран в организме человека достигает десятков тысяч квадратных метров. Мембраны выполняют несколько важных функций.
Первая – барьерная. Она обеспечивает селективный, регулируемый, пассивный и активный обмен веществ клетки с окружающей средой. Селективный – значит избирательный. Одни вещества переносятся через биологические мембраны, другие нет. Регулируемый – проницаемость мембраны для определенных веществ зависит от функционального состояния клетки. Активный – перенос от мест, где концентрация вещества мала, к местам с большей концентрацией.
Вторая – матричная. Она обеспечивает взаимное расположение мембранных белков и их оптимальное взаимодействие (например, взаимодействие мембранных фрагментов). И, наконец, третья – механическая. Эта функция обеспечивает прочность и автономность клеток и внутриклеточных структур.
Кроме того, биологические мембраны, выполняют и другие весьма важные функции: энергетическую (синтез АТФ на внутренних мембранах митохондрий и фотосинтез углеводов в мембранах хлоропластов), генерацию и проведение биопотенциалов и рецепторную (механическая, акустическая, обонятельная, зрительная, химическая).
Мембрана создает существенные ограничения для проникновения различных веществ, однако она не является полностью непроницаемой. Небольшие нейтральные молекулы могут проникать через бислой в области «структурных» дефектов. Целостная структура мембраны создается за счет гидрофобных и электростатических взаимодействий, а не за счет ковалентных связей между составляющими ее молекулами белков и липидов. Гидрофобный липидный бислой представляет естественную преграду для проникновения полярных молекул. Мембраны асимметричны по своему исходному строению, что обеспечивает градиент кривизны и спонтанное образование замкнутых структур. Мембраны животных содержат три основных компонента: фосфолипиды, гликолопиды и холестерол. Несмотря на различия в составе, все мембранные липиды построены по единому плану и легко смешиваются друг с другом, образуя монослойные или бислойные структуры (рис. 6.12). В этих структурах реализуется два типа взаимодействий: ионные взаимодействия полярных «головок» и гидрофобные взаимодействия жирнокислотных цепей. Благодаря этому мицеллы и липосомы, создаваемые протяженными бислойными структурами, достаточно стабильны в водном окружении.
Отличительной чертой процесса образования липидного бислоя является способность к самоорганизации, поскольку подобные структуры оказываются устойчивыми не за счет ковалентных связей между образующих их молекулами, а по термодинамическим причинам. Формирующаяся в результате подобных процессов мембрана состоит из двойного слоя липидов с двумя гидрофильными поверхностями (экстрацеллюлярной и протоплазматической) и гидрофобной сердцевины (рис. 6.12). При этом для образования участка мембраны площадью 1 мкм2 требуется около 5×106 липидных молекул.
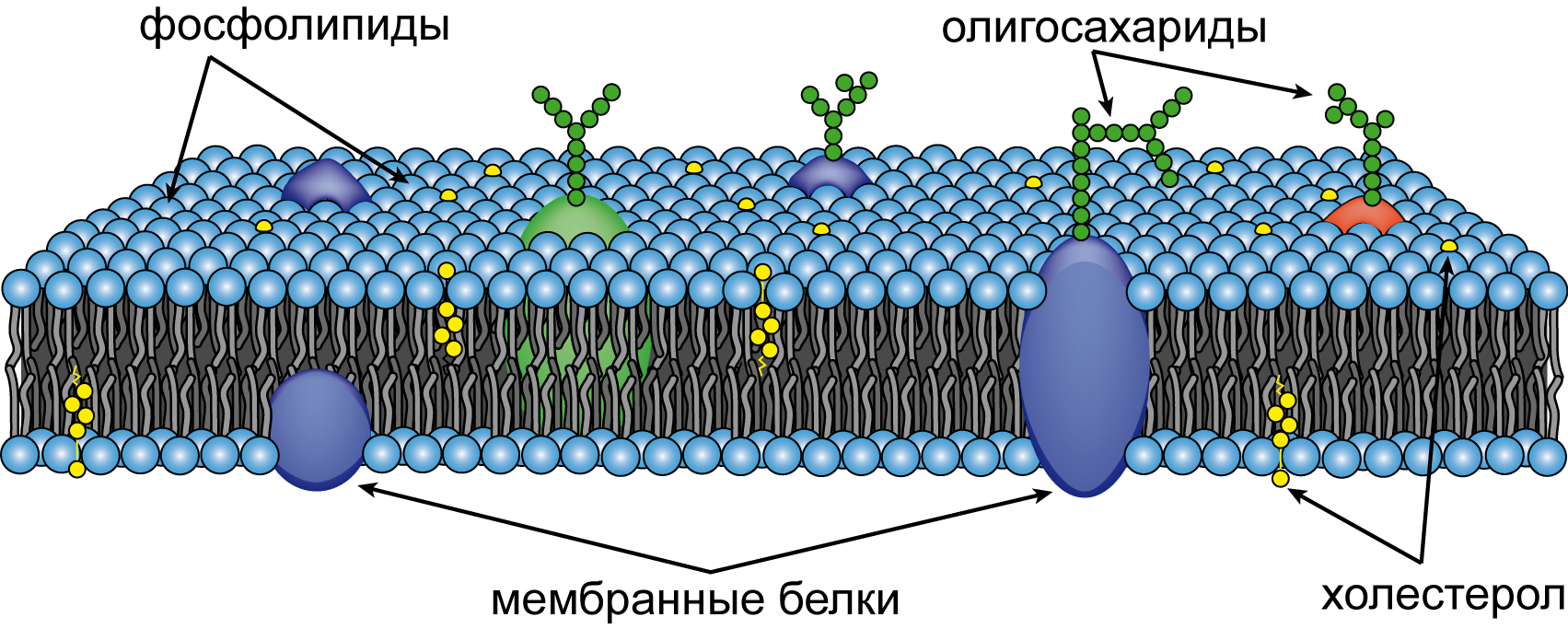
Рис. 6.12. Схематическое представление строения клеточной мембраны
Такая мембрана обладает универсальной особенностью – текучестью, под которой понимают нахождение мембранных липидов в состоянии непрерывного движения, связанного как с их вращением вокруг собственной оси, перпендикулярного плоскости бислоя (вращательная диффузия), так и с боковыми перемещениями в плоскости занимаемого ими слоя мембраны (латеральная или боковая диффузия). Обсуждение такой диффузионной подвижности, как с теоретической, так и экспериментальной точки зрения будет дано в Лекции 22.
Хотя основные структурные особенности цитоплазматической мембраны определяются свойствами липидного бислоя, большинство ее специфических функций осуществляется белками, которые составляют приблизительно 50% от массы большинства клеточных мембран и находятся в ней в соотношении одна белковая молекула на пятьдесят молекул липидов. Существует три основных способа ассоциации мембранных белков с липидным бислоем. Белки первого типа называются трансмембранными. Они пронизывают мембрану насквозь в виде одиночной или нескольких α-спиралей, и погружены в нее почти перпедикулярно. Типичным примером такого белка является родопсин. Белковые молекулы второго типа называются интегральными белками. Они погружены в гидрофобную область только одного липидного слоя и часто оказываются связанными с формирующими его молекулами ковалентными связями. Так, принимающий участие в процессах электронного транспорта цитохром с расположен с наружной стороны мембраны, в то время как F1-фрагмент АТФ-синтазы располагается на ее внутренней стороне. Белки третьего типа, называемые периферическими, в отличие от трансмембранных и интегральных белков не погружены в липидный бислой и не соединены с ним ковалентно. Они удерживаются вблизи мембраны электростатическими взаимодействиями. Описание свойств мембранных белков и их функций выходит за рамки нашей лекции, однако к структуре мембранных белков мы вернемся в Лекции 70, посвященной методу ЯМР.
Долгое время для описания цитоплазматических мембран использовалась модель неструктурированной двумерной жидкости, в которую встроены индивидуальные белки, Эта модель получила название жидкой мозаичной модели. Позже она была заменена на более структурированную модель, в которой постулировано существование кластеров мембранных белков, имеющих размеры от нескольких до сотен нанометров. Поэтому вопрос о диффузионной подвижности таких кластеров является одной из ключевых задачей мембранологии. Его рассмотрение будет дано в Лекциях 18 и 19, посвященных поступательному и вращательному трению, соответственно. В заключение заметим, что большинство мембранных белков способны вращаться вокруг своей оси или перемещаться латерально в плоскости мембраны и вносят свой вклад в ее текучесть. Однако как мы покажем в Лекции 22, скорости такой диффузии гораздо меньше, чем у мембранных липидов.