Бічні шлуночки
Бічні шлуночки (правий та лівий) є порожнинами кінцевого мозку. Вони складаються із переднього, заднього, нижнього рогів та центральної частини (рис. 5.31). Передній ріг знаходиться у лобовій частці, задній - у потиличній, нижній - у скроневій, а центральна частина - у тім'яній. Стінки рогів утворені білою речовиною півкуль та хвостатим ядром. Шлуночок заповнений спинномозковою рідиною і через міжшлуночковий отвір сполучається із III шлуночком.
Біла речовина півкуль
Біла речовина півкуль займає простір між корою і базальними ядрами. Вона складається з численних нервових волокон, що йдуть у різних напрямках. Виділяють три системи волокон півкуль; асоціативні (з'єднують різні ділянки однієї півкулі), комісуральні (з'єднують симетричні ділянки лівої та правої півкуль) і проекційні (з'єднують кору півкуль з іншими відділами головного мозку та спинним мозком). Останні отримали назву провідних шляхів.
Спинномозкова рідина: загальні дані, практичне значення
Спинномозкова рідина (lіqиоr сеrеbrоsріпаlis) міститься у підпавутинному просторі, шлуночках і центральному каналі спинного мозку. У дорослої людини кількість спинномозкової рідини становить 120-150 мл, вона виробляється судинними сплетеннями шлуночків головного мозку. Спинномозкова рідина циркулює у всіх напрямках. З бокових шлуночків вона надходить через міжшлуночкові отвори у порожнину третього шлуночка, а потім (через водопровід середнього мозку) у порожнину четвертого. Звідси через серединний (Мажанді) та бічний (Люшке) отвори частина спинномозкової рідини потрапляє у підпавутинний простір головного та спинного мозку, частина її по центральному каналу спинного мозку поширюється у каудальному напрямку, у розширення підпавутинного простору в ділянці кінського хвоста. Далі вона потрапляє у підпавутинний простір спинного мозку і рухається вгору до головного мозку. Відтік (всмоктування) спинномозкової рідини відбувається у венозну систему через грануляції павутинної оболонки.
Функції спинномозкової рідини різноманітні: вона є амортизатором для головного та спинного мозку, що оберігає його від механічних ушкоджень; бере участь у живленні та метаболічних процесах нервової тканини; у захисті від водно-осмотичних зрушень, забезпечує нормальний кровообіг у порожнині черепа.
В спинномозковій рідині знаходяться фізіологічне активні речовини, які можуть дифундувати до нервових центрів, змінюючи їх активність. Через цей механізм опосередковується регуляторна функція ліквору.
У клінічній практиці велике значення має вивчення її при різних захворюваннях нервової системи, а також можливість ендолюмбального введення лікарських речовин, обминаючи гематоенцефалічний бар'єр як для його зміцнення, так і для безпосереднього впливу на патологічний процес.
Найчастіше ліквор одержують за допомогою поперекової (люмбальної) пункції, яку проводять звичайно між II і IV або IV і V поперековими хребцями (щоб не пошкодити речовину спинного мозку).
Пройшовши міжостьові зв'язки та тверду мозкову оболонку, голка потрапляє у підпавутинний простір. Для діагностичних цілей беруть близько 10 мл рідини, у якій визначають тиск, колір, клітинний склад, хімічні складники (електроліти, білок, глюкозу), виявляють збудників захворювань.
Отже, розглянемо топографію деяких кіркових кінців різних аналізаторів (ядер) відносно часток і звивин півкуль великого мозку у людини.
1. У корі зацентральної звивини знаходиться ядро кіркового аналізатора загальної чутливості (больової, температурної, тактильної) і пропріоцептивної. Причому, у верхній її частині знаходиться проекція нижніх кінцівок і тулуба, нижче - верхніх кінцівок і зовсім внизу - голови. Аферентні імпульси від тактильних, температурних, больових і пропріорецепторів направляються сюди у складі висхідних чутливих шляхів. Переключення цих трактів відбувається на рівні специфічних ядер таламуса.
Головна функція сенсорної зони кори полягає в інтегративній критичній оцінці чутливої інформації, яка надходить з таламуса. Кора оцінює інтенсивність відчуттів, визначає просторові взаємозв'язки ділянок тіла, що подразнюються.
О скільки
висхідні чутливі шляхи перехрещуються
на шляху до кори, то при
ураженні кори однієї з півкуль порушується
чутливість протилежної половини
тіла.
скільки
висхідні чутливі шляхи перехрещуються
на шляху до кори, то при
ураженні кори однієї з півкуль порушується
чутливість протилежної половини
тіла.
2. Ядро рухового аналізатора розташоване в передцентральній закрутці. Звідси від пірамідних клітин Беца починається кірково-спинномозковий шлях. На рівні довгастого мозку цей руховий тракт перехрещується і закінчується на мотонейронах передніх рогів спинного мозку, а потім уже по їхніх волокнах надходить до скелетних м'язів.
Внаслідок перехресту цього тракту при ураженні однієї з півкуль головного мозку настає парез (часткове випадання рухів) або параліч (повна відсутність рухів) протилежної половини тіла. Морфо-функціональна організація рухового центру кори має свої особливості. Центри різних м'язових груп представлені неоднаково і займають нерівномірні ділянки. Найбільша регуляторна ділянка у звивині відведена м'язам голови (язик, голосовий апарат, мімічні м'язи) і кисті руки (рис. 5.29).
Ядро аналізатора співдружнього повороту голови та очей в протилежний бік залягає у задніх відділах середньої лобової звивини.
Ядро рухового аналізатора письмових знаків ("центр письма") розташоване у задніх відділах середньої лобової звивини.
Ядро рухового аналізатора артикуляції мови (центр Брока) знаходиться у задніх відділах нижньої лобової звивини зліва.
Ядро аналізатора стереогнозії (упізнавання предметів на дотик) залягає у верхній тім'яній часточці.
Ядро рухового аналізатора цілеспрямованих координованих рухів, набутих у процесі трудової діяльності (центр праксії) знаходиться в надкрайовій закрутці.
Ядро зорового аналізатора письмових знаків (центр читання) розташоване в кутовій звивині.
Ядро кіркового аналізатора зору залягає по "берегах" острогової борозни у язиковій звивині та клині. В цю зону надходить зорова інформація від фото рецепторів (паличок та колбочок) сітківки у складі зорових трактів.
У людини порушення зорової ділянки однієї з півкуль викликає випадання зору кожного ока. Якщо уражена зорова ділянка лівої півкулі, то випадають функції носової частини сітківки правого і скроневої частини сітківки лівого ока. Така особливість пояснюється частковим перехрестом зорового тракту: перехрещуються лише медіальні волокна тракту, а латеральні не перехрещуються. Таким чином, у кожну півкулю поступає зорова інформація від обох очей.
10. Ядро кіркового аналізатора слуху знаходиться у звивині Гешля (середній відділ на медіальній поверхні верхньої скроневої звивини). У цьому кірковому центрі звукові сигнали, що приходять із завитки внутрішнього вуха по слухових шляхах, сприймаються як звуки, що відрізняються за тоном, якістю і гучністю.
11. Ядро слухового аналізатора мовних сигналів знаходиться у задніх відділах верхньої скроневої звивини (центр Верніке).
12. Ядро кіркового аналізатора нюху та смаку розташоване у гачку, зубчастій звивині, гіпокампі.
2.4 - 2.5. Структурно-функціональна організація автономної нервової системи. Симпатичний, парасимпатичний та метасимпатичний відділи, їхня роль у регуляції вісцеральних функцій.
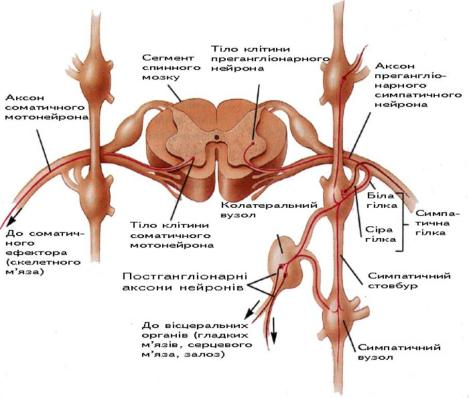
Як вивчали раніше, єдина нервова система умовно ділиться на дві частини: соматичну (анімальну) та вегетативну. Вегетативна (автономна) нервова система складається з двох частин: симпатичної і парасимпатичної, вона іннервує всі органи і тканини: залози, гладкі м'язи судин та внутрішніх органів, серцевий м'яз, органи відчуття, а також головний і спинний мозок (рис. 5.51).
Більшість органів іннервуються як симпатичним, так і парасимпатичним відділами вегетативної нервової системи. Парасимпатичний відділ не іннервує посмуговані м'язи, гладкі м'язи матки, більшість кровоносних судин, сечоводи, потові залози, волосяні фолікули шкіри, селезінку, надниркові залози та гіпофіз.
Вегетативна нервова система не має своїх особливих аферентних, чутливих шляхів. Чутливі імпульси від органів йдуть у складі чутливих волокон, які є спільними для вегетативної та соматичної нервових систем. Вищий контроль та регуляція функцій вегетативної нервової системи, як і соматичної, відбувається за рахунок кори півкуль великого мозку.
Соматична та вегетативна нервові системи відрізняються не лише за функцією, а й за будовою, а саме:
1. Соматичні рухові волокна виходять із спинного мозку сегментарно, а вегетативні волокна виходять лише з певних центрів: мезенцефалічного, бульбарного (парасимпатичні ядра черепних нервів у ромбоподібній ямці), які об'єднані в краніальний відділ; тораколюмбального (бічні роги протягом сегментів від СVIII до LII-III), сакрального (бічні роги протягом сегментів S II-IV).
Тораколюмбальний відділ належить до симпатичного відділу, а краніальний і сакральний - до парасимпатичного.
Соматичні нервові волокна мають добре помітну м'якотну оболонку, а у вегетативних вона відсутня.
Тіла клітин соматичних нейронів знаходяться у передніх рогах спинного мозку, а їхні відростки (аксони), не перериваючись, досягають скелетних м'язів. Нервові відростки вегетативних нейронів перериваються у вегетативних вузлах. Отже, тут є передвузлові (прегангліонарні) і післявузлові (постгангліонарні) волокна.
Найпростіша рефлекторна дуга соматичної нервової системи складається з двох нейронів, а вегетативної - з трьох (є вставний нейрон); її еферентна ланка складається не з одного, а з двох нейронів.
Функції організму поділяють на соматичні і вегетативні. До соматичних відносять чуттєве сприйняття подразнень і рухові реакції, що здійснюються скелетними м'язами. До вегетативних - функції, від яких залежить обмін речовин в організмі (кровообіг, дихання, травлення, виділення і т.ін.), а також ріст і розмноження.
Соматична нервова система забезпечує сенсорні і моторні функції організму, а вегетативна - регуляцію діяльності внутрішніх органів, потових залоз, обміну речовин. Таким чином, вегетативна нервова система підтримує гомеостаз.
К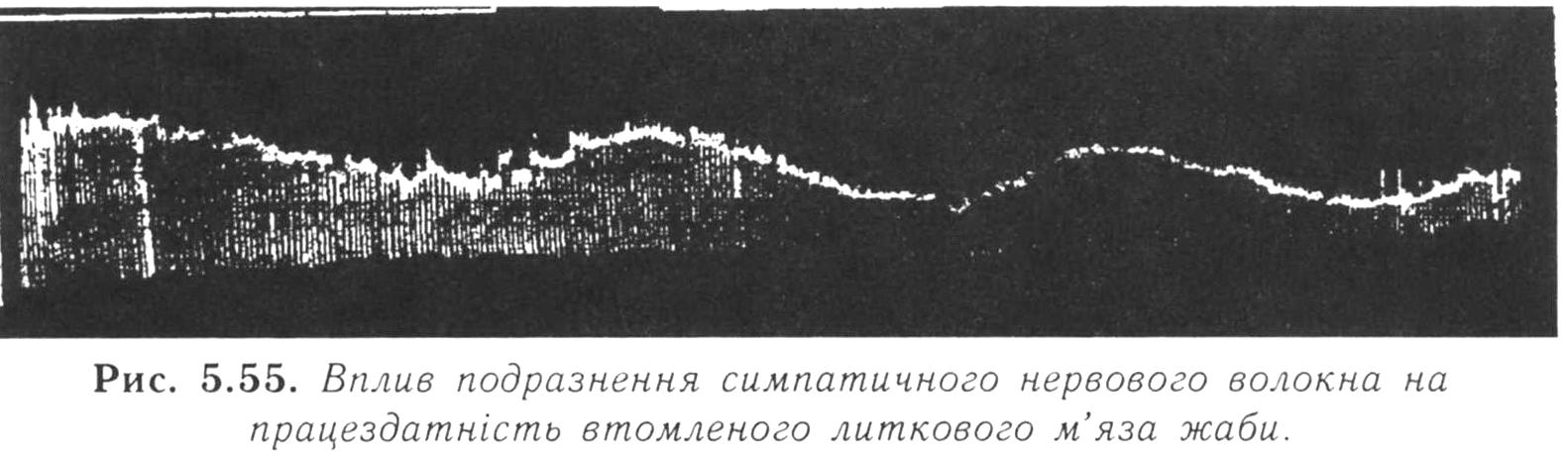 рім
цього, вегетативна нервова система
виконує адаптаційно-трофічну роль.
Це показали Л.А. Орбелі та А.Г. Пнецинський
у експерименті на нервово-м'язовому
препараті жаби (рис. 5.55). Вони подразнювали
електричним струмом руховий
нерв, при цьому м'яз скорочувався до
розвитку втоми. Після цього до подразнення
рухового нерва приєднали подразнення
симпатичного нерва, який-теж
іннервує цей м'яз. При цьому м'яз
відновлював свою працездатність і знову
починав
скорочуватись.
рім
цього, вегетативна нервова система
виконує адаптаційно-трофічну роль.
Це показали Л.А. Орбелі та А.Г. Пнецинський
у експерименті на нервово-м'язовому
препараті жаби (рис. 5.55). Вони подразнювали
електричним струмом руховий
нерв, при цьому м'яз скорочувався до
розвитку втоми. Після цього до подразнення
рухового нерва приєднали подразнення
симпатичного нерва, який-теж
іннервує цей м'яз. При цьому м'яз
відновлював свою працездатність і знову
починав
скорочуватись.
Відповідно до теорії Орбелі - Тінецинського, симпатична нервова система виконує трофічну функцію, яка полягає у збільшенні обмінних процесів у тканинах і підвищенні їх працездатності.
Адаптаційний (пристосувальний) вплив симпатичної нервової системи полягає в тому, що під її дією організм готується до роботи в нових, змінених умовах.
Існує взаємозв'язок між емоційним станом і активністю симпатичної нервової системи. Стан гніву, люті або, навпаки, радості викликає підвищення діяльності симпатичної нервової системи і, як наслідок - викид адреналіну мозковим шаром надниркових залоз у кров. Відбувається активація так званої симпатоадреналової системи, яка діє як єдине ціле.
У більшості органів, що іннервуються вегетативною нервовою системою, подразнення симпатичних і парасимпатичних волокон викликає протилежний ефект.
При подразненні симпатичних нервів поліпшується кровообіг, звужуються кровоносні судини (крім судин серця, мозку і скелетних м'язів, що працюють), зростає рівень артеріального тиску, розширюються бронхи, збільшується частота і глибина дихання, зростає легенева вентиляція, покращується постачання систем організму поживними речовинами. Симпатична частина підвищує обмін речовин, енергії та рівень глюкози в крові. Симпатичні нерви гальмують секрецію, моторику і всмоктування в шлунково-кишковому тракті і викликають розширення зіниці ока. Діяльність симпатичної частини збільшує резервні можливості кожної з функціональних систем.
При подразненні парасимпатичних нервів зменшується сила і частота серцевих скорочень, розширюються судини, падає артеріальний тиск, дихання стає нечастим і поверхневим, звужуються бронхи, зменшується легенева вентиляція, звужується зіниця. З боку шлунково-кишкового тракту збільшується секреція, моторика і всмоктування в ньому, розслаблюються сфінктери і скорочується мускулатура травного каналу. Парасимпатична частина зменшує обмін речовин і енергії, знижує рівень глюкози в крові.
Незважаючи на протилежні ефекти симпатичної і парасимпатичної частин вегетативної нервової системи, вони являють собою єдине ціле. В регуляції функцій цілісного організму важливою є діяльність як однієї, так і другої частини. Тільки узгоджена діяльність обох частин вегетативної нервової системи зумовлює нормальне функціонування організму.
Метасимпатический отдел вегетативной нервной системы
Кроме парасимпатического и симпатического отделов, физиологи выделяют метасимпатический отдел вегетативной нервной системы. Под этим термином подразумевается комплекс микроганглионарных образований, расположенных в стенках вн>тренних органов, обладающих моторной активностью (сердце, кишечник, мочеточники и др.) и обеспечивающих их автономию. Функция нервных узлон заключается В передаче центральных (симпатических, парасимпатических) влияний к тканям, и, кроме того, они обеспечивают интеграцию информации, поступающей по местным рефлекторным дугам. Метасимпати-ческие структуры представляют собой самостоятельные образования, способные функционировать при полной децентрализии. Несколько (5—7) из относящихся к ним близлежащих узлов объединяются в единый функциональный модуль, основными единицами которого являются клетки-осцилляторы, обеспечивающие автономию системы, интернейроны, мотонейроны, чувствительные клетки. Отдельные функциональные модули составляют сплетение, благодаря которому, например, в кишке организуется перистальтическая волна.
Функции метасимпатического отдела вегетативной нервной системы напрямую не зависят от деятельности симпатической или парасимпатической
нервных систем, но могут видоизменяться под их влиянием. Так, например, активация парасимпатического влияния усиливает перистальтику кишечника, а симпатического — ослабляет ее.
МЕТАСИМПАТИЧНА НЕРВОВА СИСТЕМА
Слід зазначити, що це поняття «молоде», у підручниках анатомії, гiстологii, фiзiогii його не згадують. Ввів цей термін А. Д. Ноздрачев. Він вважає, що метасимпатична нервова система (МНС) - це комплекс мiкроганглiонарних утворів, розташованих у стінках внутрішніх органів, що володіють моторною активністю. Мова йде про «наявність мiкроганглiiв (iнтрамуральних ганглiiв) у шлунку, кишечнику, сечовому мiхуру, серцi, бронхах. З погляду органної приналежності мiкроганглiiв А. Д. Ноздрачев пропонує виділити відповідно кардiометасимпатичну, ентерометасимпатичну, уретрометасимпатичну, везикулометасимпатичну нервову систему. В матцi, в області її шийки, теж є метасимпатична система. Найбільше вивчена метасимпатична система кишечника і серця.
Було давно відомо, що в ШКТ є нервові сплетення - пiдсерозне, мiжм`язеве (Ауербахове) і подслизисте (Мейсснерово). У кожному з цих сплетень є множина мiкроганглiiв, в яких виділяють 3 типа нейронів (по Догелю). 1-й тип нейронів по Догелю являють собою еферентнi нейрони, аксон яких безпосередньо контактує з м'язевою клітиною. Нейрони II типу по Догелю являють собою аферентнi нейрони. Їх аксони можуть переключатися на нейрони 1-го типу (рефлекторна дуга замикається на рівні мiкроганглiя), або аксон може йти до паравертебральному або превертебральному ганглiям, переключаючись тут на інші нейрони, або нейрони, або аксони цих аферентних нейронів можуть доходити до спинного мозку і тут переключатися на інші нейрони. Т. е. аферентна iмпульсацiя, що йде від мiкроганглiiв, може замикатися на різних рівнях. Нейрони 3 типу по Догелю являють собою асоціативні нейрони.
Аналогічна картина характерна і для метасимпатичних структур серця. Яку ж функцію і яким чином здійснює метасимпатична нервова система? Метасимпатична система може, по-перше, здійснювати передачу центральних впливів - за рахунок того, що парасимпатичнi і симпатичні волокна можуть контактувати з метасимпатичною системою і тим самим корегувати її вплив на об'єкти керування. По-друге, метасимпатична система може виконувати роль самостійного утворення, що інтегрує, тому що в ній є готовірефлекторні дуги (аферентнi - вставні - еферентнi нейрони).
Т. И. Косицький (разом з іншими фізіологами) показав, що в ізольованому
серці має місце процес рефлекторної регуляції: розтяг правого передсердя збільшує роботу правого желудочка серця. Цей ефект блокується ганглiоблокаторами. Аналогічно - розтяг правого шлуночка серця підвищує роботу лівого шлуночка. Реакція гальмується ганглiоблокаторами.
В ШКТ метасимпатична нервова система здійснює регуляцію складних рухiв кишки - перистальтику, маятникоподiбнi рухi. Це складний процес, у котрому багато ще залишається неясним. Думають, що завдяки рефлекторним дугам, що починаються з рецепторів (хемо-, механо-), можлива тонка регуляція моторики кишечника, адаптована до процесів гiдролiзу і всмоктування поживних речовин у шлунково-кишковому тракті.
Детальне вивчення мікроструктури і функціональної організації
мiкроганглiiв ШКТ дозволило А. Д. Ноздрачеву сформулювати уявлення про те, що в основі діяльності метасимпатичноi нервової системи лежить функціональний модуль: це скупчення певним чином пов'язаних між собою нейронів, що і забезпечують функцію метасимпатичноi системи. У цьому модулі виділяють клiтини-осцилятори, сенсорні нейрони, мотонейрони і iнтернейрони. Ключовою клітиною модуля є клiтина-осцилятор. Вона спонтанно збуджується у певному ритмі, і її потенціали дії передаються
через систему вставних нейронів до мотонейрона, тобто руховому нейрону, аксони якого контактують з м'язевою клітиною. Якщо мова йде про модуль ШКТ, то в даному випадку контакт здійснюється з ГМК кишечника. Медіатором мотонейрона є АТФ (нейрон - пуринергiчний), що блокує автоматичну активність ШКТ. Таким чином, чим активніше
клiтина-осцилятор, тим виражене гальмування, що здійснює мотонейрон на ГМК кишечника. Вся ця система «осцилятор - мотонейрон» піддається модуляції з декількох сторін: а) аферентнi нейрони, збуджуючись в результаті активації їх закінчень (рецепторів), можуть змінювати активність мотонейрона, діючи на нього безпосередньо (активація через холінергічний синапс) або на його закінчення (аксо-аксональне
гальмування), знімаючи гальмiвний вплив на ГМК; б) парасимпатичнi і симпатичні постгангліонарнi волокна, дiючи на вставні нейрони,- модулюють стан мотонейрона. Наприклад, у ШКТ при активації парасимпатичних волокон відбувається гальмування мотонейрона метасимпатичного модуля, що знімає гальмування з боку метасимпатичного
модуля на ГМК, і в результаті - активація ГМК. Симпатичнi волокна, навпаки, посилюють гальмiвний вплив пуринергічного мотонейрона на ГМК шлунка або кишечника.
Отже, серед нейронів метасимпатичної нервової системи є пуринергічнi,
холiнергічнi нейрони, а також (про них ще не говорилося) адренергiчнi,
серотонiн-ергiчнi і, можливо, гiстамiнергiчнi нейрони.
2.6 – 2.8. Структурно-функціональна організація ендокринної системи. Ендокринні залози, їхні гормони та значення. Основні механізми дії гормонів. Регуляція секреції гормонів. Механізм взаємодії ендокринних залоз. Гіпотоламо-гіпофізарна система. Роль ліберинів і статинів. Аденогіпофіз, його гормони, механізм впливу. Щитоподібна залоза, її гормони та вплив на обмін речовин. Прищитоподібні залози, їхні гормони та функції. Ендокринна функція підшлункової залози. Поняття про стрес. Роль гормонів надниркових залоз під час стресу.
Залозами внутрішньої Секреції, або ендокринними, називаються такі залози, які не мають вивідних проток і секрет (інкрет, гормон) яких надходить безпосередньо у кров або лімфу.
Процес синтезу і виділення у внутрішнє середовище фізіологічно активних речовин ендокринними залозами називають внутрішньою секрецією, а ці речовини - гормонами.
До ендокринних залоз відносяться: гіпофіз, епіфіз, прищитоподібні, щитоподібна, загруднинна, надниркові залози. Підшлункова та статеві залози є змішаними, в них виробляються і гормони, і продукти зовнішньої секреції.
Крім ендокринних залоз, існують паракринні залози і клітини, які розміщені в органах (шлунок, сечівник, нирки) серед інших тканин і секретують тканинні гормони.
Найбільш поширеною класифікацією залоз внутрішньої секреції вважають класифікацію за Альошиним, згідно з якою всі ендокринні залози поділяють на чотири групи: 1) аденогіпофіз і залежні від нього ендокринні залози (щитоподібна, яєчко, яєчники, кірковий шар надниркових залоз); 2) незалежні від аденогіпофіза (паращитоподібні, загруднинна, мозкова речовина надниркових залоз, панкреатичні острівці підшлункової залози); 3) "нервового походження" (гіпоталамус, кишкові та шлункові клітини); 4) нейрогліального походження (епіфіз, нейрогіпофіз).
Залежно від походження із різних видів епітелію ендокринні органи поділяють на п'ять груп (Заварзін О.О., Щелкунов С.І.).
Залози ентодермального походження, які розвиваються із епітеліальної вистилки глоткової кишки - так звана бранхіогенна група. Це щитоподібна та прищитоподібні залози.
Залози ентодермального походження - із епітелію кишкової трубки - ендокринна частина підшлункової залози (панкреатичні острівці).
Залози мезодермального походження - інтерреналова система, кіркова речовина надниркових залоз та інтерстиціальні клітини статевих залоз.
Залози ектодермального походження - похідні переднього відділу нервової трубки (неврогенна група) - гіпофіз та шишкоподібне тіло (епіфіз мозку).
5. Залози ектодермального походження - похідні симпатичної частини вегетативної нервової системи. До цієї групи належать мозкова речовина над ниркових залоз та параганглії.
Гормони - це фізіологічне активні речовини, які синтезуються спеціалізованими клітинами ендокринних залоз. Вони надходять безпосередньо в кров і проявляють свою дію лише на складні клітинні структури - клітинні мембрани, ферментні системи.
За хімічною структурою розрізняють стероїдні гормони (кортикостероїди, андрогени, естрогени), білкові (інсулін і глюкагон), похідні тирозину (тироксин, норадреналін), похідні жирних кислот (простагландини).
Гормони опосередковують свої дії через рецептори органів-мішеней. Розрізняють два типи гормональної рецепції: внутрішньоклітинний і поверхневий мембранний. У першому випадку гормони легко проникають у клітину через мембрану і зв'язуються з рецептором у цитоплазмі, а далі комплекс гормон-рецептор транспортується у ядро і вступає у взаємодію з хроматином. При цьому активується синтез РНК, моделюються процеси транскрипції ДНК і РНК, проходить довготривала перебудова клітинного метаболізму.
Другий шлях рецепції - це зв'язок гормонів з рецепторами на поверхні клітини. При цьому відбувається утворення із АТФ гормонального посередника - циклічного 3,5-аденозинмонофосфату. Останній діє на ферменти клітини і таким чином змінює її метаболізм та проникність.
Впливаючи на синтез білка в клітинах, гормони регулюють фізичний, статевий і розумовий розвиток, обмін речовин і енергії, сприяють підтриманню гомеостазу організму (рівня глюкози, електролітів, осмотичного тиску, артеріального тиску), забезпечують адаптацію активності фізіологічних систем. Вони впливають на поведінку і психіку людини. Під дією гормонів проходять зміни не тільки фізіологічних процесів, а й морфологічних.
Синергізм - це дія в одному напрямку. З точки зору кібернетики - це прояв принципу дублювання, надлишковості, яка забезпечує високу надійність у підтримуванні гомеостазу. Наприклад, гормон мозкового шару надниркових
залоз адреналін і гормон підшлункової залози глюкагон активують розпад глікогену печінки до глюкози і викликають підвищений рівень цукру в крові.
Антагонізм - це дія в протилежних напрямках. Прикладом може бути гормон підшлункової залози інсулін, який знижує рівень цукру в крові, і гормон кори надниркових залоз адреналін, який підвищує рівень цукру. Але діючи антагоністично, ці гормони в кінцевому результаті сприяють покращанню вуглеводного живлення тканин.
Суть пермисивної дії полягає в тому, що гормон, не викликаючи фізіологічного ефекту, створює умови для реакції органа на дію другого гормону. Наприклад, глюкокортикоїди не впливають на тонус судин, однак створюють умови, при яких надпорогові концентрації адреналіну підвищують артеріальний тиск.
Механізм роботи гіпоталамо-гіпофізарно-адреналової системи, її роль в адаптації організму. Ендокринні, паракринні залози.
Діяльність ендокринних залоз регулюється нервовою системою. Є два шляхи такої регуляції. По-перше, через гіпоталамо-гіпофізарну систему: нейросекреторні клітини гіпоталамуса продукують рилізинг-гормони. Останні потрапляють у передню частку гіпофіза через його ворітну систему вен і стимулюють (ліберини) чи гальмують (статини) вивільнення його гормонів. Ці гормони надходять у загальний кровотік, досягають щитоподібної залози, кори надниркових залоз і сприяють секреції гормонів цими залозами.
По-друге, центральна нервова система через еферентні впливи вегетативної нервової системи регулює діяльність підшлункової залози, мозкового шару надниркових залоз.
Регуляція функції ендокринних залоз, як і будь-яка регуляція, має принцип саморегулювання, який відбувається на основі зворотного зв'язку. Це забезпечує стабілізацію активності залоз внутрішньої секреції. Наприклад, збільшення рівня цукру в крові призводить до підвищення концентрації інсуліну.