

Орієнтовні рефлекси
Орієнтовні рефлекси ще називають сторожовими, або, за влучним висловом І. П. Павлова, рефлексами «що таке?» Вони полягають у швидкому повороті голови, тулуба, очних яблук у напрямку світлового або звукового подразника. У рефлексах беруть участь передні горбки чотиригорбкового тіла (первинні зорові центри) — у відповідь на світлове подразнення виникають зорові орієнтовні рефлекси, а також задні (первинні центри слуху) — у відповідь на звукові подразники формуються слухові орієнтовні рефлекси.
Завдяки цим рефлексам організм орієнтується у нових ситуаціях, готується до захисту, нападу чи інших дій відповідно до світлових або звукових подразнень. Одночасно відбувається перерозподіл тонусу м'язів — переважно підвищується тонус флексо-рів, посилюється діяльність серця та ін. Ушкодження горбиків чотиригорбкового тіла призводить до втрати здатності визначати джерела світлових чи звукових подразників. Через первинні зорові центри замикається ряд інших рефлексів — відбувається акомодація та конвергенція очей.
Значення червоних ядер
Від червоних ядер середнього мозку починається руброспінальний шлях, який зразу після початку переходить на протилежний бік. Шлях цей іде в бокових стовпах спинного мозку, його волокна закінчуються переважно на інтернейронах, а частина — на мотонейронах. Основний вплив червоних ядер полягає в збудженні α- та γ-мотонейронів-згиначів і в гальмуванні мотонейронів-розгиначів.
У червоному ядрі існує певне топографічне представництво скелетних м'язів. Локальне подразнення окремих його відділів викликає скорочення одного м'яза чи груп м'язів.
Руброспінальний шлях функціонує в тісному взаємозв'язку з пірамідним шляхом. Вони обидва йдуть у латеральних стовпах, впливають переважно на функції дистальних груп м'язів, і тому їх називають латеральною руховою системою (на відміну від вестибулоретикулоспінальної медіальної рухової системи).
Участь проміжного мозку в регуляції рухових функцій
До проміжного мозку належать таламус і гіпоталамус. До складу моторних ядер таламусу входить вентролатеральне ядро. Саме сюди спрямовуються імпульси від мозочка, базальних гангліїв, а від цих ядер — до моторної зони кори великого мозку.
Гіпоталамічна ділянка має численні аферентні та еферентні зв'язки з різними відділами ЦНС і грає головну роль у підтриманні гомеостазу, впливаючи на функції вегетативної нервової та ендокринної систем. Гіпоталамус бере участь у регуляції обміну речовин, функцій органів травлення та кровообігу, підтриманні температури тіла на постійному рівні, а також у формуванні багатьох поведінкових реакцій, пов'язаних із голодом, насиченням, спрагою, сном, статевими реакціями тощо. Важлива роль гіпоталамусу в пристосуванні (адаптації) організму до різних умов існування, зокрема до фізичної праці.
Роль переднього мозку в регуляції рухових функцій
До переднього мозку належать базальні ганглії (СПРС № 4) та кора великого мозку.
Роль кори великого мозку в регуляції рухових функцій
Кора великого мозку — філогенетично наймолодша структура мозку. В процесі еволюції ссавців особливо швидко розвивалась нова кора, її товщина досягла 1—2 мм, а загальна її поверхня у людини становить близько 2200 см2. До складу кори великого мозку входять нейрони, які в різних її ділянках мають свої особливості. Розрізняють понад 50 цитоархітектурних полів. Багато з них беруть участь у регуляції рухових функцій.
Сенсорні зони кори великого мозку тісно пов'язані з моторними зонами, які лежать перед центральною борозною, займаючи приблизно задню частину лобної частки. У моторній корі розрізняють: первинну, премоторну та додаткову моторну ділянки.
Первинна
моторна кора
міститься перед центральною борозною
(поле 4). Як і перша соматосенсорна зона,
ділянка має топічну організацію.
Подразнення електричним струмом верхньої
частини
ділянки у тварин чи у людей під час
нейрохірургічних операцій
супроводжується скороченням м'язів
нижньої частини тіла на протилежному
боці. Понад 50 % поверхні цієї ділянки
у людини — це представництво рук та
мови, що свідчить про розвиток цих
функцій у людини (мал. 93.).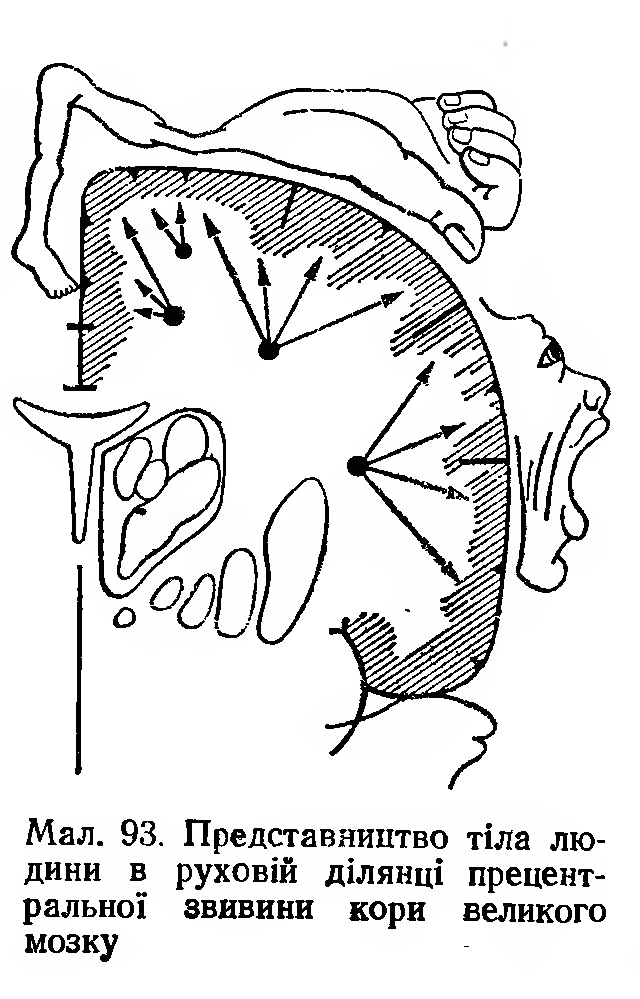
Премоторна зона лежить попереду первинної моторної (поле 6) і в глибині сільвієвої борозни. Більшість імпульсів із цієї зони викликає моделі рухів, вмикаються групи м'язів, які виконують певні функції. Імпульси йдуть у первинну моторну кору, до базальних ядер, а потім знову в моторну зону через таламус. Ця зона разом із базальними ядрами, таламусом, первинною моторною корою керує багатьма складними рухами. Додаткова моторна ділянка розташована перед і над премоторною зоною. Щоб' скоротились м'язи, контрольовані цією зоною, треба застосувати електричний Струм значно більшої потужності. Ефект подразнення часто виявляється з обох боків тіла. Функція розглядуваної ділянки полягає в тому, що вона разом з премоторною ділянкою створює умови для забезпечення пози і рухів, що реалізуються вже первинною моторною корою. Нейрони кіркової моторної зони одержують численні імпульси волокнами, що йдуть від сенсомоторної ділянки парієтальної кори, зорової і слухової ділянок, моторної кори протилежного боку, таламуса, а також від базальних ядер та інших структур.
Кортико-спінальний (пірамідний) шлях розпочинається від нейронів моторної кори, проходить між хвостатим ядром та шкаралупою (внутрішня капсула). У довгастому мозку більшість волокон (80%) переходить на протилежний бік — у латеральний пірамідний шлях, а частина (20 %) —на протилежний бік нижче (у шийному та грудному відділах спинного мозку), тобто у вентральний пірамідний шлях.
У пірамідному шляху міститься близько 1 млн волокон. Лише незначна частина їх (близько 3 %) починається від гігантських пірамідних клітин Беца. Колатералі від волокон пірамідного шляху йдуть у сусідні ділянки кори великого мозку і зумовлюють гальмування (таким чином запобігають іррадіації збудження). Більшість волокон латерального пірамідного шляху закінчується на інтернейронах спинного мозку, частина — на сенсорних нейронах задніх рогів або власне на мотонейронах.
Роль пірамідного шляху в здійсненні рухової функції полягає в регуляції тонких і точних рухів, особливо дистальних частин кінцівок, тобто пальців. При порушенні кровопостачання моторної кори (крововилив, тромбоз, травма, пухлина) спостерігається параліч м'язів на протилежному боці тіла. Значна кількість волокон від моторної кори не йде безпосередньо до спинного мозку пірамідним шляхом, а перемикається на різних ядрах — базальних, червоному, вестибулярних, ядрах ретикулярної формації та ін.