

Додаток до лекції № 2 ( до 5 практичної роботи). Роль заднього мозку в регуляції рухових функцій
До цього відділу ЦНС належать довгастий мозок та міст головного мозку. Основні функції заднього мозку в регуляції рухових функцій: 1) аналіз сенсорної інформації, що надходить в ядра заднього мозку від рецепторів шкіри та м'язів голови, і еферентна іннервація скелетних м'язів голови;
2) аналіз сенсорної інформації, що надходить від вестибулярних рецепторів, пропріорецепторів м'язів шиї, тулуба, кінцівок, та рефлекторна регуляція тонусу м'язів тіла, яка забезпечує певну позу (тонічні рефлекси пози);
3) провідникова функція — проходження нервових імпульсів, що йдуть у вищерозташовані відділи ЦНС та від них до скелетних м'язів.
Сенсорна інформація надходить від рецепторів шкіри обличчя, слизових оболонок носа, зубів, окістя кісток черепа, пропріорецепторів окорухових м'язів, смакових рецепторів, рецепторів вестибулярного апарата та слуху, глотки, рецепторів багатьох внутрішніх органів.
Еферентна іннервація забезпечує функцію жувальних, окорухових, мімічних м'язів, м'язів язика, глотки, гортані, шиї. Ці функції забезпечуються нервами, що ще зберігають здатність до сегментарної іннервації, але вона виражена менш чітко, ніж у спинному мозку.
Н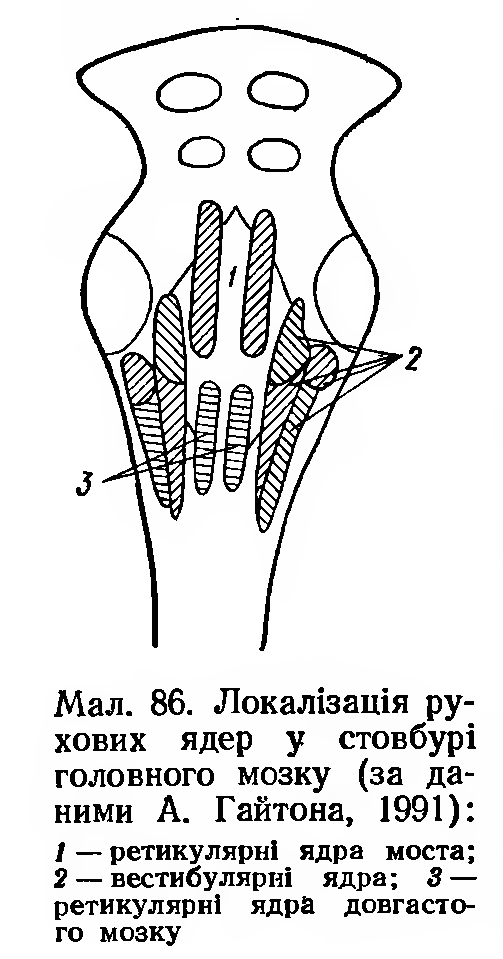 адзвичайно
велику роль відіграє задній мозок у
підтриманні пози антигравітації
(забезпечується тонусом м'язів) та
перерозподілі тонусу при зміні
положення голови. Жаба, у якої перерізаний
мозок між довгастим та спинним (так
звана спінальна тварина), не може
підтримувати тонус скелетних м'язів
на такому рівні, якого було б досить для
створення пози антигравітації. Така
тварина лежить розпластана на
горизонтальній поверхні.
адзвичайно
велику роль відіграє задній мозок у
підтриманні пози антигравітації
(забезпечується тонусом м'язів) та
перерозподілі тонусу при зміні
положення голови. Жаба, у якої перерізаний
мозок між довгастим та спинним (так
звана спінальна тварина), не може
підтримувати тонус скелетних м'язів
на такому рівні, якого було б досить для
створення пози антигравітації. Така
тварина лежить розпластана на
горизонтальній поверхні.
Жаба, у якої перерізаний мозок вище від заднього, має позу, яка дозволяє перебороти силу гравітації Землі. Для створення такої пози важливе значення має тонус м'язів-екстензорів. Цей тонус має рефлекторну природу, бо він зникає після перерізування задніх корінців нервів, що входять у спинний чи довгастий мозок. Центрами регуляції є три рухові ядра заднього мозку: 1) вестибулярне ядро; 2) ядра ретикулярної формації варолієвого моста; 3) ядра ретикулярної формації довгастого мозку (мал. 86).
За участю цих центрів реалізуються тонічні рефлекси пози, серед яких розрізняють вестибулярні статичні та шийні.
Вестибулярні статичні рефлекси розпочинаються з рецепторів переддвер'я — сферичного та еліптичного мішечків. Вони були детально досліджені голландським фізіологом К. Мадпіїй (1924).
Існує кілька видів вестибулярних статичних рефлексів: 1) при підніманні голови підвищується тонус м'язів-екстензорів передніх кінцівок (мал. 87) і знижується тонус цих м'язів задніх кінцівок;
2) при нахилянні голови вниз спостерігається протилежний ефект;
3) при нахилянні голови вбік збільшується тонус м'язів-екстензорів з того боку, куди нахилена голова.
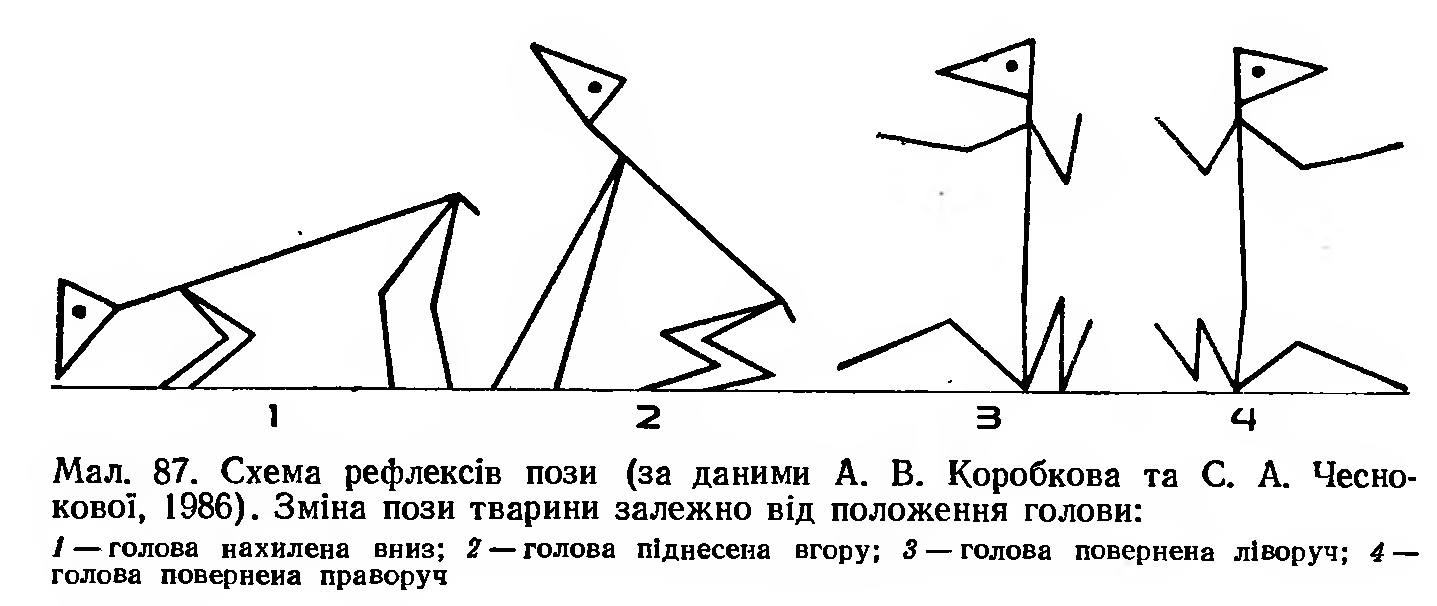
Значення цих рефлексів полягає в тому, що відбувається перерозподіл тонусу м'язів при зміні положення голови, і це забезпечує підтримання рівноваги. Волокна вестибулоспінального шляху йдуть своїм боком часто моносинаптично до α- та γ-мотонейронів передніх рогів спинного мозку. Під впливом цих волокон збуджуються α- та γ-мотонейрони екстензорів і гальмуються мотонейрони флексорів. При порушенні вестибулярного апарата чи вестибулярних ядер спостерігаються істотні зміни в регуляції тонусу скелет' них м'язів, а у зв'язку з цим порушення і в локомоціях.
Шийні тонічні рефлекси дублюють вестибулярні статичні рефлекси. Ці рефлекси починаються з пропріорецепторів м'язів шиї. Імпульси надходять у рухові центри довгастого мозку, а звідти — до мотонейронів спинного мозку. Ці рефлекси впливають не лише на м'язи кінцівок, а й на очні м'язи (забезпечується компенсаторна установка очних яблук при зміні положення голови). Відхилення голови супроводжується рухом очних яблук упротилежному напрямку. Таким чином зберігається правильна зорова орієнтація. У чистому вигляді шийні рефлекси можна спостерігати після руйнування вестибулярного апарата.
Вимкнути шийні рефлекси можна за допомогою перерізування задніх корінців відповідних сегментів спинного мозку або накладання гіпсової пов'язки на шию, яка робить нерухомими шийні м'язи.
Вплив ретикулярної формації стовбура головного мозку на діяльність рухових центрів спинного був відкритий І. М. Сєченовим у 1862 р. Механізм гальмування рефлексів спинного мозку можна пояснити постсинаптичним гальмуванням мотонейронів, впливом на проміжні інтернейрони або на терміналі аферентних волокон, які входять у спинний мозок. Зараз відомо, що локальне подразнення гігантоклітинного ядра ретикулярної формації довгастого мозку зумовлює гальмування згинальних та розгинальних рефлексів спинного мозку.
Крім гальмівних впливів інші ділянки ретикулярної формації (латеральні зони) можуть полегшувати спінальні рефлекси. При цьому знижується поріг та скорочується латентний період цих рефлексів.
Вплив ретикулярної формації здійснюється через два ретику-лоспінальні шляхи — медіальний та латеральний. Медіальний шлях розпочинається від варолієвого моста і, не перехрещуючись, йде до сегментів спинного мозку. При цьому збуджуються мото-нейрони екстензорів і гальмуються мотонейрони флексорів. Волокна цього шляху проходять у передніх відділах спинного мозку поруч із вестибулоспінальним шляхом і зумовлюють подібний ефект.
Ретикулярна формація довгастого мозку діє протилежним чином. Вона збуджує α- та γ-мотонейрони флексорів і гальмує мотонейрони екстензорів.