

3 Структура и состав биомембран
Мембраны состоят из липидов трёх классов: фосфолипиды, гликолипиды и холестерол. Фосфолипиды и гликолипиды (липиды с присоединёнными к ним углеводами) состоят из двух длинных гидрофобных углеводородных «хвостов», которые связаны с заряженной гидрофильной «головой». Холестерол придаёт мембране жёсткость, занимая свободное пространство между гидрофобными хвостами липидов и не позволяя им изгибаться. Поэтому мембраны с малым содержанием холестерола более гибкие, а с большим — более жёсткие и хрупкие. Также холестерол служит «стопором», препятствующим перемещению полярных молекул из клетки и в клетку. Важную часть мембраны составляют белки, пронизывающие её и отвечающие за разнообразные свойства мембран. Их состав и ориентация в разных мембранах различаются.
Клеточные мембраны часто асимметричны, то есть слои отличаются по составу липидов, переход отдельной молекулы из одного слоя в другой (так называемый флип-флоп) затруднён.
Биомембраны: структура и функции
А. Структура плазматической мембраны
Все биомембраны построены одинаково; они состоят из двух слоев липидных молекул толщиной около 6 нм, в которые встроены белки. Некоторые мембраны содержат, кроме того, углеводы, связанные с липидами и белками. Соотношение липиды : белки : углеводы является характерным для клетки или мембраны и существенно варьирует в зависимости от типа клеток или мембран.
Компоненты мембран удерживаются нековалентными связями, вследствие чего они обладают лишь относительной подвижностью, т. е. могут диффундировать в пределах липидного бислоя. Текучесть мембран зависит от липидного состава и температуры окружающей среды. С увеличением содержания ненасыщенных жирных кислот текучесть возрастает, так как наличие двойных связей способствует нарушению полукристаллической мембранной структуры. Подвижными являются и мембранные белки. Если белки не закреплены в мембране, они «плавают» в липидном бислое как в жидкости. Поэтому говорят, что биомембраны имеют жидкостно-мозаичную структуру.
В то время как «дрейф» в плоскости мембраны происходит достаточно легко, переход белков с внешней стороны мембраны на внутреннюю («флип-флоп») невозможен, а переход липидов происходит крайне редко. Для «перескока» липидов необходимы специальные белки транслокаторы. Исключение составляет холестерин, который может легко переходить с одной стороны мембраны на другую.
Б. Мембранные липиды
На рисунке схематически изображена биомембрана. В мембранах содержатся липиды трех классов: фосфолипиды, холестерин и гликолипиды. Наиболее важная группа, фосфолипиды, включает фосфатидилхолин (лецитин), фосфатидилэтаноламин, фосфатидилсерин, фосфатидилинозит и сфингомиелин. Холестерин присутствует во внутриклеточных мембранах животных клеток (за исключением внутренней мембраны митохондрий). Гликолипиды входят в состав многих мембран (например, во внешний слой плазматических мембран). В состав гликолипидов входят углеводные функциональные группы, которые ориентируются в водную фазу.
Липиды мембран представляют собой амфифильные молекулы с полярной гидрофильной головкой (голубого цвета) и неполярным липофильным хвостом (желтого цвета). В водной среде они агрегируют за счет гидрофобных взаимодействий и вандерваальсовых сил.
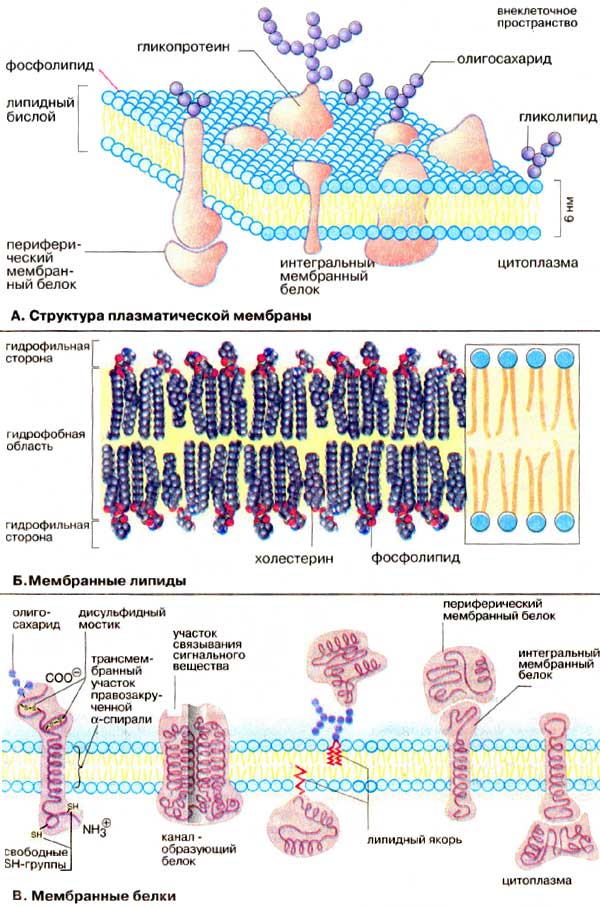
В. Мембранные белки
Протеины могут связываться с мембраной различным путем.
Интегральные мембранные белки имеют трансмембранные спирализованные участки (домены), которые однократно или многократно пересекают липидный бислой. Такие белки прочно связаны с липидным окружением.
Периферические мембранные белки удерживаются на мембране с помощью липидного «якоря» и связаны с другими компонентами мембраны; например, они часто бывают ассоциированы с интегральными мембранными белками.
У интегральных мембранных белков фрагмент пептидной цепи, пересекающий липидный бислой, обычно состоит из 21-25 преимущественно гидрофобных аминокислот, которые образуют правую α-спираль с 6 или 7 витками (трансмембранная спираль).
Дополнительная информация
Белки клеточной поверхности и некоторые липидные молекулы несут ковалентно связанные углеводные компоненты, экспонированные на наружной стороне мембраны. Эти гликопротеины и гликолипиды вместе с дополнительными несвязанными гликопротеинами и полисахаридами образуют клеточную оболочку (гликокаликс).
СТРУКТУРА И ФУНКЦИИ БИОМЕМБРАН
Биомембраны характеризуются чрезвычайным разнообразием и способны не только отделять содержимое клетки от внешней среды и обеспечивать разделение внутреннего объема клетки на компартменты, но и участвовать в регуляции множества процессов. Например, плазматические мембраны обеспечивают диффузионный барьер, активный транспорт, электрическую возбудимость, межклеточную коммуникацию, гормональный и иммунный ответы и др. На мембранах эндоплазматической сети происходит синтез белков, жиров и углеводов. Мембраны нервных клеток способны передавать импульсы в форме изменения электрического потенциала и т.д. Уникальность функций каждой мембраны в значительной степени определяется свойствами мембранных белков, входящих в ее состав.
Белки — разнообразные ферменты, транспортные белки, рецепторы, поры, каналы и др. — вносят существенный вклад в формирование структуры клеточных мембран. Среднее содержание белков в мембранах составляет примерно 60% (по массе сухого вещества), при этом в состав биомембран также входят липиды — 30% и углеводы — 10%. Естественно, соотношение между этими компонентами может значительно меняться в зависимости от природы мембран. Так, содержание белков в мембранах может варьировать от 20% в миелине до 80% в митохондриях. Липиды — фосфолипиды, гликолипиды, холестерин — составляют костяк мембраны и ответственны за целостность мембранной структуры. Углеводы обнаруживаются в составе мембранных белков (гликопротеинов и протеогликанов) или липидов (гликолипидов). Кроме того, в мембранах содержится относительно большое количество (~30%) связанной невымерзающей воды.
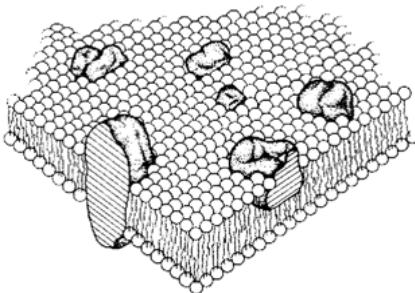
Несмотря на многообразие биомембран основные принципы структурной организации всех мембран животного, растительного и бактериального происхождения одинаковы. Согласно получившей широкое признание «жидкостно-мозаичной» модели, первоначально предложенной в 1972 году Сингером и Николсоном, биомембрана представляется как текучий фосфолипидный бислой, в который погружены белки (рис. 1 а). Впоследствии стало, однако, очевидным,
Рис. 1. Модели структурной организации биологических мембран.
(а) — «жидкостно-мозаичная» модель, в рамках которой мембрана представляется как текучий фосфолипидный бислой, в который погружены свободно диффундирующие белки.
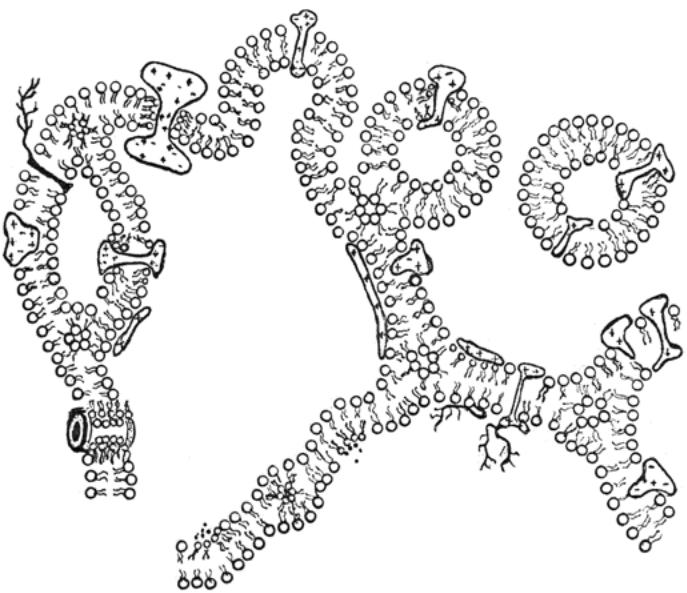
(б) — «метаморфно&мозаичная» модель, включающая в себя следующие процессы:
1. Трансмембранный перенос полярных молекул и ионов (Са2+ и т.д.), связанный с возникновением промежуточных образований типа обращенных мицелл;
2. Сочленение мембран друг с другом;
3. Слияние мембран и экзоцитоз;
4. Трансмембранный перенос белка;
5. Компартментализация в протяженных мембранных системах;
6. Образование пор в мембране.
что молекулярная организация мембран гораздо сложнее, чем это следует из жидкомозаичной модели. В частности, показано, что не все мембранные белки свободно диффундируют в жидком липидном бислое. Некоторые участки мембран отличаются по своей структуре от классического липидного бислоя вследствие липидного полиморфизма. В пределах одной мембраны могут соседствовать участки с разным липидным составом и функциями. В настоящее время считают, что сложная динамическая структура биомембран, для которой характерны искривления, фазовые переходы, вариации толщины, образование небислойных структур, определяется специфическими взаимодействиями мембранных белков с липидами. Такие взаимодействия во многом обеспечивают эффективное выполнение мембранами разнообразных клееточных функций, возникающих в ходе метаболизма. Проиллюстрировать динамические свойства биомембран можно на примере «метаморфно&мозаичной» модели, включающей в себя основные мембранные процессы (рис. 1 б).
 а)
а)
 б)
б)