

2. Нейрофизиологические механизмы регуляции бодрствования
Задание 3: Составьте сравнительную характеристику основных различий в эффектах активации ствола мозга и таламуса на основе материала учебника Основы психофизиологии / Под ред. Ю.И. Александрова. М., 2008. (с. 156). Данные занесите в таблицу:
Характеристики реакции |
ЭЭГ активация ствола |
ЭЭГ активация таламуса |
Область распространения |
Генерализированная |
Локальная |
Тип реакции |
Тоническая |
Фазическая |
Временная динамика |
Медленно угасающие |
Быстро угасающие
|
|
|
|
Задание 2: Рассмотрите рисунок, выделите регуляторные блоки функциональных состояний. Используя знания физиологии ЦНС, расскажите об уровнях регуляции бодрствования: клеточном, отдельных центров и мозга как целого.
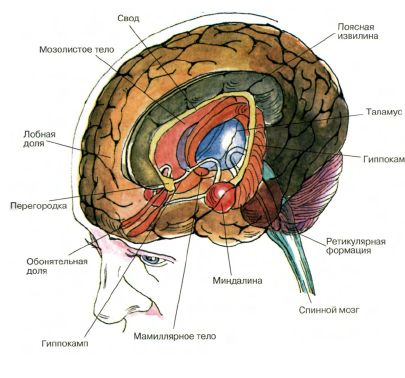
Регуляторные блоки функциональных состояний: 1)Ретикулярная формация,2) Передние отделы коры больших полушарий — фронтальные зоны,3)Таламус,4)Кора больших полушарий,5) поясная извилина, 6)гиппокамп.
Клеточный уровень регуляции бодрствования: На нейронном уровне регуляция функциональных состояний осуществляется с помощью особой категории нейронов, именуемых модуляторными. Существуют две категории модуляторных нейронов: активирующего и инактивирующего типа. Первые увеличивают активность синапсов, соединяющих чувствительные и исполнительные нейроны, вторые снижают эффективность синапсов, прерывая путь передачи информации от афферентных к эфферентным нейронам. Кроме того, нейроны-модуляторы различаются по степени генерализованности своего действия. Переход к бессознательному состоянию, например при засыпании, можно определить как выключение активирующих нейронов-модуляторов генерализованного типа и включение инактивирующих нейронов-модуляторов. В эволюции нейроны-модуляторы объединились в ансамбли и сети, сосредоточенные на уровне ретикулярной формации ствола мозга и неспецифического таламуса, образуя активирующую и инактивирующую системы.
Ур.отдельных центров: Совокупность модулирующих систем образует особый блок, которая регулирует тонус коры и подкорковых структур, оптимизирует уровень бодрствования в отношении выполняемой деятельности и обуславливает адекватный выбор поведения в соответствии с актуализированной потребностью. Важнейший участок регуляторного блока — ретикулярная формация мозга, представляющая сеть из нервных клеток в средней части ствола. С боковых сторон ретикулярная формация окружена сенсорными путями, которые отдают часть афферентной импульсации в ретикулярную формацию. Благодаря этому любое сенсорное возбуждение повышает уровень активации ретикулярной формации, активация по восходящим путям распространяется вверх к коре больших полушарий. Экспериментально показано, что раздражение ретикулярной формации через вживленные в нее электроды приводит к пробуждению спящего животного.Еще одно важное звено регуляции функциональных состояний связано с работой таламуса. Зрительный бугор, или таламус — отдел промежуточного мозга, который выполняет роль коллектора сенсорной информации, так как в него поступает информация от всех органов чувств. По некоторым данным, в центре таламуса находится "водитель ритма" — морфо-функциональное образование, отвечающее за генерацию ритмической активности и распространяющее синхронизированные влияния на обширные области коры. Ядра неспецифического таламуса образуют диффузную проекционную таламическую систему, которая оказывает на кору возбуждающие и тормозные влияния. Эти влияния по сравнению с эффектами возбуждения ствола мозга имеют более ограниченный характер и захватывают относительно небольшие участки коры.Таким образом, при раздражении таламуса возникает реакция активации в коре головного мозга. Эта реакция отчетливо видна в текущей электроэнцефалограмме: она сравнительно кратковременна и локализована. В отличие от реакции активации, вызываемой ретикулярной формацией ствола мозга, которая считается генерализованной реакцией активации, эффекты возбуждения неспецифического таламуса называют локальной активацией.
Ур.мозга как целого: Важнейшим регулятором уровня бодрствования в целом и внимания как избирательного процесса служат передние отделы коры больших полушарий - фронтальные зоны. Именно эти структуры по нисходящим кортикоретикулярным путям модулируют в нужном направлении активность стволовой и таламической систем. Включение в этот процесс фронтальных зон с их нисходящими путями позволяет говорить о существовании своеобразного замкнутого контура регуляции. Исходно ретикуляторная формация ствола мозга, возбуждаясь под действием внешних стимулов, активизирует неспецифический таламус и кору больших полушарий, а та в свою очередь, благодаря нисходящим проводящим путям может снизить активность ретикулярной формации ствола и таламуса или увеличить в зависимости от того, что требуется в данный момент времени. Таким образом, можно говорить о существовании регулируемой или управляемой корковой активации, за счет которой кора больших полушарий может настраивать собственный уровень возбудимости соответственно задачам текущей жизнедеятельности.