

12.3. Фізіологія слуху
Звук — коливання повітряного чи водного середовища або твердого субстрату — відіграє у житті багатьох тварин подвійну роль. З одного боку, це сигнал про небезпеку з боку природного явища чи ворога, а з другого, — це спосіб спілкування особин одного виду, якщо ці особини мають звукоутворювальні органи.
12.3.1. ЕВОЛЮЦІЯ СЛУХУ
Переважна більшість тварин сприймає звук за допомогою спеціалізованих органів слуху. У риб слухову рецепцію частково здійснює також бічна лінія.
Слух у безхребетних тварин. Звуко-сириймальиі органи з'явились на досить ранніх етапах еволюції тваринного світу. Так, вже у медуз функцію слухової рецепції викопують статоцисти, які сприймають інфразвукові коливання в межах 8 — 13 Гц. У молюсків цей діапазон становить від кількох герц до 2 кГц, хоча самі молюски звуків не генерують. Сприймання і відтворення звуків добре розвинуті у членистоногих, зокрема комах. Комахи сприймають звукові коливання за допомогою волоскових сенсил, джоистоиових і тимпанальних органів, які розміщені в різних місцях: па церках, антенах, голові, черевці, кінцівках. У головоногих молюсків і примітивних хордових слухова рецепція не розвинена.
Слух у хребетних тварин. Орган слуху у всіх хребетних, починаючи з пай-примітивніших круглоротих, знаходиться у лабіринті внутрішнього вуха, точніше, в одному з мішечків лабіринту. Розміщені па стінках мішечка скупчення волоскових сенсорних клітин (плями) у цих тварин поряд з гравітаційними силами сприймають і звукові коливання. У риб від мішечка відходить тяж — лагена (попередник завитки ссавців), яка бере па себе звукосириймальиу функцію. Оскільки зовнішнього і середнього вуха з барабанною перетинкою у риб немає, роль резонатора, що підсилює звук, частково бере на себе плавальний міхур, який сполучається з лагепою за допомогою чотирьох веберових кісточок. Завдяки добре розвиненому апарату веберових кісточок Карпові риби чують звуки чи не найкраще серед риб. Діапазон звуків, які вони сприймають, зростає до 4 —6 кГц. До того ж вони здатні розрізняти 1/3 — 1/4 музичного тону.
Для наземних тварин у повітряному середовищі звук відіграє значно більшу роль, ніж для водних організмів. Тому у них відбувається прогресивний розвиток всієї слухової системи: у амфібій з'являється барабанна перетинка, у рептилій —зовнішній слуховий хід. Слуховий стовпчик (колумелла) у них з'єднує барабанну перетинку із внутрішнім вухом, але у змій вій прикріплюється не до барабанної перетинки, а до квадратної кістки черепа, що робить їх дуже чутливими до коливань ґрунту. Черепахи і ящірки сприймають звуки до 10 кГц, але невідомо, чи здатні вони їх розрізняти.
У деяких птахів і у всіх ссавців, за винятком китових, з'являється зовнішнє вухо, яке дає їм змогу сприймати напрям звуку. У ссавців розвивається справжня завитка, яка має від 0,25 (качкодзьоб) до 4 (свиня) обертів. Людина сприймає звуки в діапазоні 18 Гц — 20 кГц, а в середньому у класі ссавців слуховий діапазон досягає ультразвукових частот — 50 Гц — 140 кГц.
12.3.2. СЛУХОВА СИСТЕМА ССАВЦІВ
Орган слуху, вухо, у більшості ссавців побудоване і функціонує за однаковим принципом — виконує функції слуху і рівноваги тіла. У людини вухо складається із зовнішнього, середнього і внутрішнього вуха, де розміщений ирисінково-завит-ковий орган.
Зовнішнє вухо складається з вушної раковини і зовнішнього слухового ходу. Вушна раковина схожа на лійку, передня стійка якої зрізана, що дає змогу орієнтуватись у напрямку надходження звуку і краще сприймати звуки, що надходять
спереду. Поступове звуження вушної раковини біля переходу її у зовнішній слуховий хід дає змогу концентрувати звукові хвилі й підсилювати звук.
Зовнішній слуховий хід відділяється від барабанної порожнини середнього вуха барабанною перетинкою.
Середнє вухо. До середнього вуха належать барабанна перетинка, барабанна порожнина зі слуховими кісточками (молоточок, коваделко, стремінце) і слухова (євстахієва) труба, яка сполучає середнє вухо з носовою частиною глотки. За допомогою цієї слухової труби тиск у порожнині середнього вуха підтримується па рівні атмосферного, і барабанна перетинка зазнає однакового тиску з обох боків. Барабанна порожнина має шість стінок. Ла-біриитна стійка барабанної порожнини межує з внутрішнім вухом і має два отвори: верхній — вікно присінка (овальне), прикрите основою стремінця, і нижній — вікно завитки (кругле), затягнуте вторинною барабанною перетинкою.
Ланцюг слухових кісточок середнього вуха фіксується еластичними зв'язками. Крім того, у середньому вусі є два м'язи: м'яз — натягувач перетинки і стремінце-вий м'яз, які своїми скороченнями змінюють натягнення барабанної перетинки і слухових кісточок, регулюючи функцію звукопровідності.
Порожнина середнього вуха у ссавців заповнена повітрям. Слухові кісточки виконують подвійну роль. По-перше, вони утворюють систему важелів, за допомогою яких поліпшується передача звукової енергії з повітряного середовища середнього вуха до перилімфи внутрішнього вуха. Завдяки тому, що площа основи стремінця, укріпленого у вікні присінка (овальному), значно менша за площу барабанної перетинки, а також завдяки спеціальному способу сполучення кісточок, що діють як важелі, тиск па мембрану цього вікпа виявляється у 22 рази більшим, ніж па барабанну перетинку.
По-друге, коли звуковий тиск наближається до 120 дБ, характер руху слухових кісточок завдяки зміні їх натягу змінюється так, що значно знижує коефіцієнт передачі звукової енергії і запобігає виникненню больового відчуття. При цьому рефлекторне скорочення м'язів — натягувана барабанної перетинки і стреміп-цевого — під час дії звуків значної інтенсивності зумовлює зменшення амплітуди коливань барабанної перетинки, кісточок середнього вуха і відповідне зменшення звукового тиску, що передається па завитку.
Передавання звукових коливань через барабанну перетинку і слухові кісточки забезпечує повітряну провідність. Кісткова провідність забезпечує сприймання звуку, наприклад, під водою. Механізм зву-косприймапня однаковий при обох видах провідності.
Особливості будови середнього вуха у китоподібних. У дельфінів і китів вушної раковини немає. Експериментально доведено, що зовнішній слуховий хід у китоподібних не бере участі у проведенні звуку. Вважають, що хвилеводом є нижня щелепа, через яку звукова хвиля досягає структур середнього вуха, де всі три слухові кісточки жорстко з'єднані між собою.
Оскільки китоподібні ведуть водний спосіб життя, череп у них не може виконувати роль ізолятора між двома вухами. Тому у них звукоснриймальпий апарат підвішений окремо па короткій сухожил-ковій зв'язці, причому середнє і внутрішнє вуха розміщені в одному кістковому утворі. Завдяки цьому орган слуху механічно досить надійно ізольований від кісток черепа. Крім того, середнє і внутрішнє вуха оточені спеціальною пазухою, заповненою повітрям і піною з білкової емульсії, що також сприяє ізоляції слухових структур від черепа.
Внутрішнє вухо у ссавців знаходиться у піраміді скроневої кістки. Це присінко-во-завитковий орган, який складається з кісткового і перетинчастого лабіринтів. Кістковий лабіринт складається з присінка, иівколових каналів і завитки, яка власне і є органом слуху. Кістковий спіральний канал завитки — завитковий лабіринт — утворює у людини 2,5 оберту, зменшуючись у діаметрі до верхівки.
Фізіологія
слуху
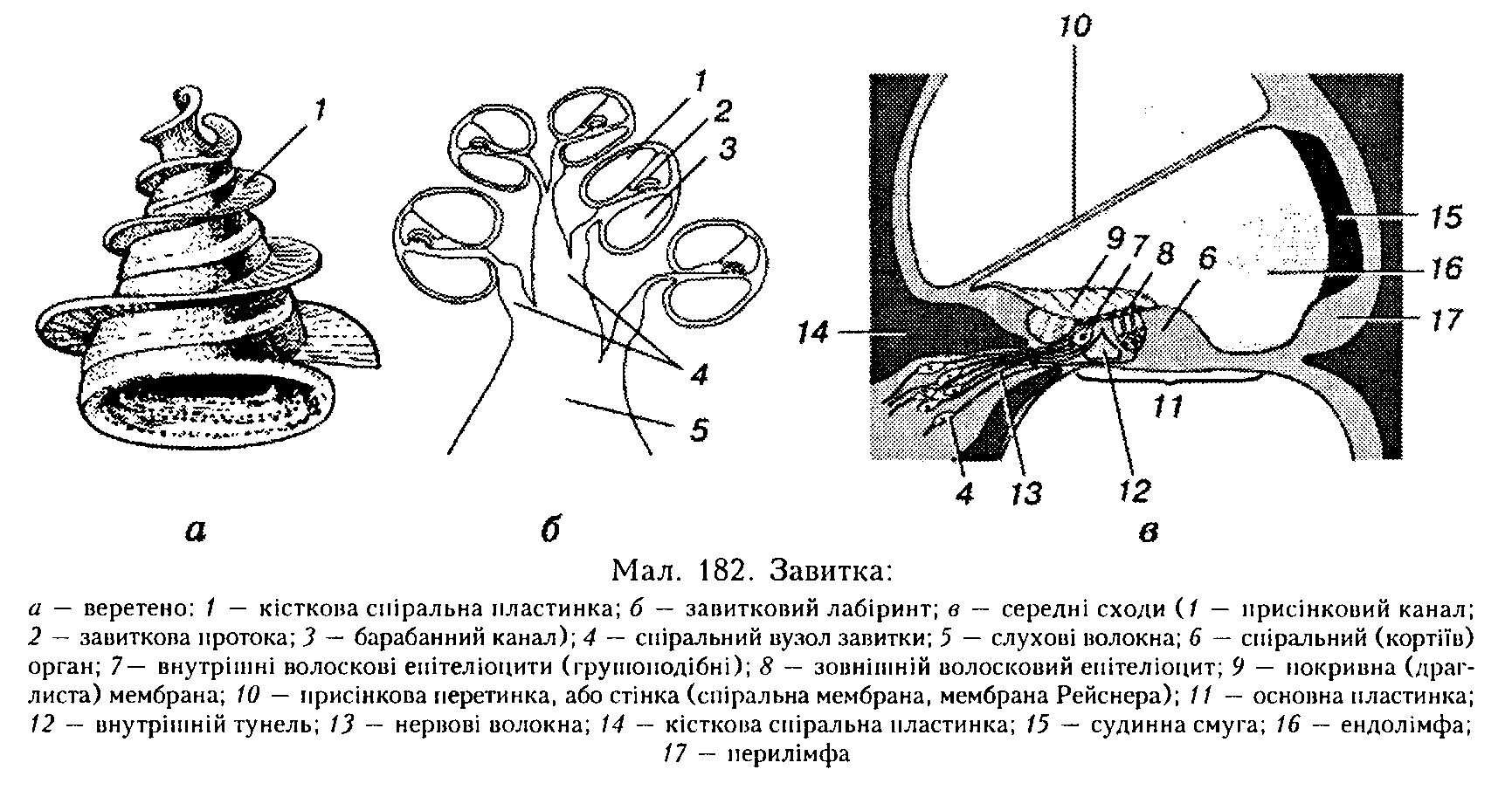
У кістковому спіральному каналі міститься перетинчаста її частина — завитковий лабіринт. Між кістковою і перетинчастою частинами завитки міститься перилімфа, а завитковий лабіринт заповнений ендолімфою. Від осі кісткового спірального каналу завитки — кісткового веретена — всередину каналу відходить уздовж всієї довжини кісткова спіральна пластинка (мал. 182, а). Вона закінчується вільним краєм, від якого до зовнішньої стінки натягнута пружна основна (базальна) пластинка (мембрана). Співвідношення між шириною кісткової пластинки і основної мембрани такі, що остання біля основи завитки є найвужчою (0,04 мм), а на верхівці завитки, навпаки, иайширшою (0,5 мм). Від кісткової спіральної пластинки відходить ще одна перетинка, яка також тягнеться до зовнішньої стінки завитки. Вона називається присінковою перетинкою, або стійкою (рейснеровою мембраною).
Мембрани ділять завитковий лабіринт па три спіральці капали, або сходи (див. мал. 182, б, в). Верхній, присінковий, канал починається від присіпкового вікна. Біля верхівки завитки він з'єднується з нижнім, барабанним, каналом через отвір завитки (гелікотрему). Середній канал — завиткова протока — закінчується біля верхівки завитки сліпим мішком. На внутрішній поверхні основної пластинки уздовж усієї довжини завиткової протоки розміщений спіральний орган.
Спіральний (кортіїв) орган (мал. 183) є звукосприймальиим рецепторним апаратом слухового аналізатора. Рецепторні клітини спірального органа — внутрішні й зовнішні волоскові сенсорні клітини — є вториипочутливими рецепторами. Внут
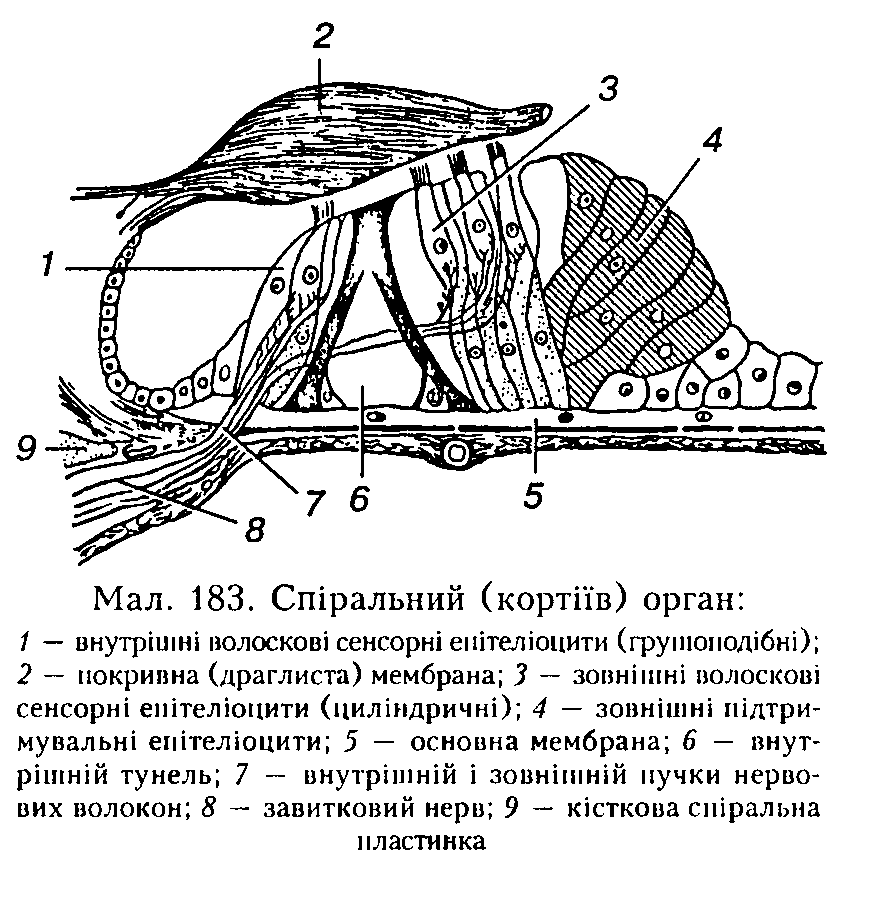
рішні волоскові клітини розміщені в один ряд, їх майже 3500. Зовнішні утворюють 3 — 4 ряди, їх значно більше — близько 12 000. Основа кожного волоскового епітеліоцита фіксується до основної мембрани, верхня частина з волосками (ЗО — 40 штук 4 — 5 мкм завдовжки— па кожній внутрішній клітині і 65 — 120 — на кожній зовнішній клітині, де вони тонші й довші) повернена до порожнини завиткової протоки.
Волоски в ендолімфі контактують з покривною (драглистою) мембраною.
Внутрішні й зовнішні волоскові еиітеліоцити мають окрему іннервацію. Волокна слухового нерва па 90 % утворюються внутрішніми пучками нервових волокон від внутрішніх волоскових епіте-л і опиті в.
12.3.3. СЛУХОВІ ПРОЦЕСИ НА РІВНІ ВНУТРІШНЬОГО ВУХА
Коли через барабанну перетинку, слухові кісточки і овальне вікно присінка звукові коливання створюють поштовхи тиску в присінку, вони поширюються в ендолімфі верхнього каналу і повертаються нижнім каналом, випинаючи перетинку круглого вікна завитки. Звукові коливання гасяться у повітряному середовищі середнього вуха, куди вони передаються через перетинку вікна завитки.
Коливання нерилімфи передаються через присіикову стінку (спіральну мембрану, мембрану Рейспера) на ендолім-фу, яка заповнює завиткову протоку, а потім — па основну мембрану, па якій розміщений спіральний орган. Під час коливань основної мембрани волоскові клітини своїми волосками дотикаються до покривної мембрани, і волоски згинаються. Така механічна деформація зумовлює деполяризацію волоскового епітеліоцита, тобто виникнення рецепторного потенціалу (РП).
Волоскові клітини є вторинпочутливи-ми рецепторами і тому їх РП спричинює появу генераторного потенціалу (ГП) у сенсорному біполярному нейроні, тіло якого знаходиться у спіральному вузлі завитки. Периферичний відросток його утворює синапси з волосковими клітинами, а центральний відросток (аксон) входить до складу слухового нерва. Заключний етап функції завитки полягає у виникненні імиульсації у волокнах завиткового нерва. Аксони нейронів завиткового вузла закінчуються па нейронах ядер довгастого мозку. До волоскових клітин підходять також і еферентні волокна, які йдуть від клітин ядра оливи довгастого мозку. Збудження цих волокон запобігає надмірному збудженню волоскових клітин.
Вухо надає інформацію про такі якості звуку, як інтенсивність і висота (частота). Якщо інформацію про силу звуку слухова система кодує і передає частотою імпульсів від кожної рецепторної клітини, то для пояснення здатності слухової системи сприймати і розрізняти звуки різної частоти А. Гельмгольц ще в XIX ст. запропонував резонансну теорію слуху. За цією теорією, кожне волокно основної мембрани має власну частоту коливань, входить у резонанс і починає коливатись, коли до завитки надходить звук, що відповідає власній частоті волокна. Разом з ним коливаються волоскові клітини спірального органа. Таким чином сприймається звук певної частоти. Проте теорія Гель-мгольця не могла пояснити цілої низки слухових явищ, тому вчені шукали нових пояснень.
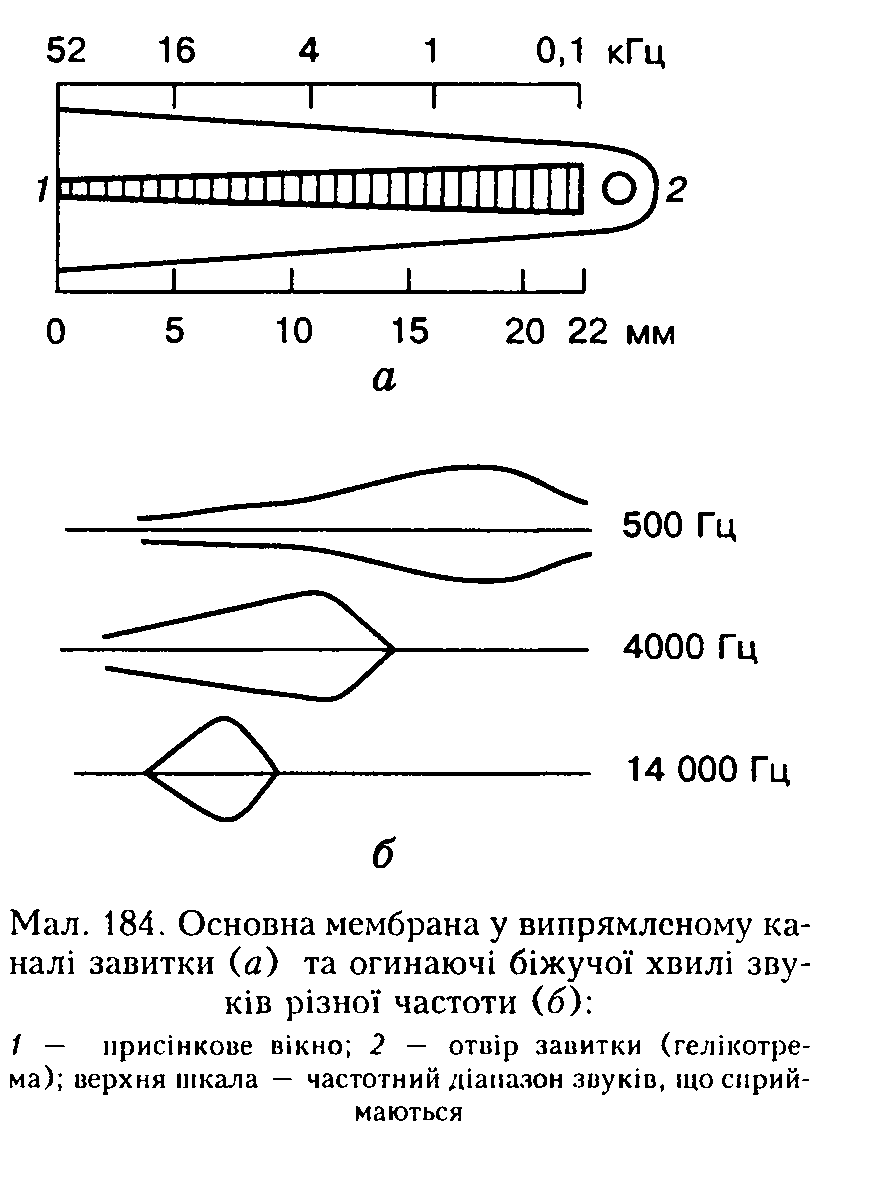
Г. Бекеші довів, що волокна основної мембрани механічно зв'язані між собою вздовж усієї довжини завиткової протоки і не можуть коливатись поодинці. Він встановив також, що основна мембрана є пай-жорсткішою біля основи завитки, тобто там, де вона пайвужча. Далі, в напрямку до верхівки, жорсткість її знижується. Саме тому під час коливання мембрани звукові хвилі поширюються від її основи до верхівки, тобто градієнт жорсткості мембрани завжди примушує хвилі рухатися від присіпкового вікна і ніколи — у зворотному напрямку. Причому високочастотні коливання поширюються по основній мембрані лише па коротку відстань, а довгі,
Фізіологія
слуху
низькочастотні, хвилі просуваються досить далеко (мал. 184). Отже, перша най-жорсткіша частина основної мембрани є високочастотним фільтром, який пропускає далі середні й довгі звукові хвилі. Така теорія звукосприймапня дістала назву теорії біжучої хвилі. Ця хвиля "біжить" вздовж основної мембрани, але її огинаюча частина стоїть нерухомо і має найбільшу амплітуду на тій ділянці мембрани, яка відповідає певній частоті хвилі.
Волоскові рецептори спірального органа збуджуються звуковою хвилею найбільше там, де амплітуда коливань максимальна, тому при дії звуку різної частоти збуджуються різні клітини (теорія місця). Біля основи завитки збудження виникає під впливом звуку високих топів, па верхівці — низьких і в середній частині — середньої частоти звукового діапазону. У ділянці максимальної амплітуди основна і покривна мембрани рухаються одна відносно іншої найбільше. Оскільки волоски рецепторних клітин утворюють щільний контакт з покривною мембраною, під час коливання основної мембрани вони згинаються, що і є початком процесу сприймання звуку.
Електричні явища. Згинання волосків рецепторних клітин викликає зміну опору їхньої мембрани. Оскільки в епдолімфі йонів калію у 100 разів більше, ніж у пери-лімфі, вона заряджена відносно иерилім-фи позитивно. Цей постійний внутрішиьо-завитковий потенціал при відсутності звуку звуку становить 80 мВ. Проте під впливом звукових коливань основної мембрани він може змінюватись. Волоскові клітини мають негативний заряд (-80 мВ), і, отже, між епдолімфою та їхнім внутрішньоклітинним середовищем створюється різниця потенціалів до 160 мВ. У зв'язку з цим навіть невеликі зміни опору мембрани волоскових клітин спричинюють виникнення локальних йонних струмів, які й зумовлюють виникнення рецепторного потенціалу.
Мікрофонний потенціал завитки. Сумарний рецепторний потенціал волоскових сенсорних епітеліоцитів відображає амплітуду і частоту звукової хвилі.
Його можна зареєструвати, якщо розташувати електроди поблизу цих клітин у барабанному каналі або прикласти їх до вікна завитки. Це й буде мікрофонний потенціал завитки. Якщо тепер його посилити і подати па гучномовець, то ми почуємо той самий звук (спів, клацання вимикачем), який діяв на вухо тварини. Це явище дістало назву мікрофонного ефекту.
12.3.4. ОБРОБКА СЛУХОВОЇ ІНФОРМАЦІЇ Б ЦНС
У наземних тварин слухові волокна — завитковий корінець присіиково-завитко-вого нерва (VIII пари черепних нервів) спрямовані до переднього і заднього завиткових ядер довгастого мозку, звідки аксони других нейронів прямують через верхню оливу до ядер трапецієподібного тіла моста, ядер нижніх горбків середнього мозку, які у нижчих хребетних тварин є вищим інтегративним центром слуху, і при
середнього колінчастого тіла проміжного мозку (метаталамус), далі до кіркового кіпця слухового аналізатора, тобто до кори верхньої скроневої завитки.
Ядра нижніх горбків середнього мозку є координатором генетично визначених рефлекторних рухів на різні характеристики звуку: його напрямок, одночасне звучання двох компонентів з різною частотою, зміни частот тощо. Особливо розвинута ця ділянка середнього мозку в кажанів і дельфінів, які використовують для своєї орієнтації і комунікації ультра-звуки.
У ссавців є п'ять основних рівнів перемикання слухових волокон: 1) завиткові ядра; 2) оливи довгастого мозку; 3) задні горбки чотиригорбкової пластинки середнього мозку; 4) ядра присереднього колінчастого тіла проміжного мозку; 5) слухова зона кори великого мозку — верхня скронева звивина. Установлено також наявність низхідних шляхів від слухової зони кори до присереднього колінчастого тіла, задніх горбків, завиткових ядер і оливи довгастого мозку. Крім того, до присереднього колінчастого тіла прямують волокна з потиличної зони кори, со-матосенсорної ділянки і ядер шатра мозочка. До задніх горбків чотиригорбкової пластинки середнього мозку підходять волокна з кількох ділянок кори великого мозку (зорової, тім'яної і рухової зон), а також від сітчастого утвору. Все це забезпечує взаємодію сенсорних систем для реалізації цілеспрямованої поведінки.
У ссавців домінує таламічний шлях аналізу звукових коливань, в якому ири-середнє колінчасте тіло метаталамуса розподіляє звукову інформацію за частотним принципом, а потім підключається слухова зона кори великого мозку, де нейрони одних колонок чутливі до звуків певної частоти, нейрони інших колонок реагують на зниження чи підвищення частоти звуків. Є також колонки, які отримують інформацію від волоскових рецепторів завитки правого і лівого вух, чутливих до тієї самої частоти, що дає змогу визначати джерело звуку у просторі. Рецепторний апарат завитки пов'язаний з різними ділянками первинної та вторинної частий слухової зони кори.
Розміри рецептивних йолів кіркових нейронів слухової зони кори досить великі, і до одного нейрона підходять волокна від рецепторів — волоскових сенсорних епітеліоцитів завитки, розміщених па різних ділянках основної мембрани завиткової протоки. Це призводить до широкого перекривання рецептивних полів нейронів з близькими частотами. Така організація рецептивних полів слухових нейронів відіграє важливу роль у механізмі кодування інтенсивності звукових подразнень. Крім того, вона забезпечує високий ступінь надійності периферичного апарату слухової системи: у разі локальних ушкоджень спірального органа виникає лише підвищення порога сприймання звуків тієї чи іншої висоти, а не повна глухота щодо цих звуків. З іншого боку, тоиотопіч-иа організація в слуховій зоні кори є нечіткою, хоча низькі тони краще представлені в задній частині слухової зони кори, тоді як високі звуки частіше аналізуються в передній її ділянці. Більшість кіркових нейронів сприймають сигнали від обох вух з переважанням сигналів від коптралате-рального вуха.
Для кожної групи рецепторних клітин існує своя оптимальна характеристична частота звуку, яка викликає найбільшу частоту розрядів ПД у волокнах завиткового нерва за найменшого порога подразнення. Вище було зазначено, що за теорією Бекеші волокна основної мембрани жорстко зв'язані між собою, і тому під впливом звуку певної частоти починає коливатись волокно, що відповідає цій частоті, а також кілька сусідніх волокон. При цьому збуджуються кілька груп волоскових сенсорних епітеліоцитів, що фіксуються до цих волокон. З посиленням звукового тиску коливний процес охоплює все більшу кількість волокон основної мембрани, активізуючи волоскові клітини з іншими характеристичними частотами. У волокнах завиткового нерва при цьому виникають ПД з частотою тим більшою, чим більше їх власна характеристична частота
Фізіологія
слуху

відповідає частоті певного звуку (мал. 185). Отже, якщо сила звукової хвилі вища за норогову, збуджуються рецептори з різними характеристичними частотами, проте ми чуємо чіткі звуки, а не безладні. Це пояснюється тим, що зменшення розмірів збудженої ділянки рецептивного поля слухової зони кори і відповідно звуження діапазону звуків, що відчуваються, забезпечується гальмуванням, і тому нейрони цього поля реагують потенціалами дії лише па тони характеристичної і близької до неї частот.
12.3.5. СПРИЙМАННЯ І РОЗРІЗНЯННЯ ЗВУКІВ
Звукові відчуття. Звук, що надходить до внутрішнього вуха, викликає певні звукові відчуття, які розрізняються за гучністю (силою звуку) і тональністю (частотою звукових коливань). Крім меж діапазону зазначених параметрів звуку показником слухової системи тварин є також здатність розрізняти звуки за силою і висотою (амплітудпо-частотпий розрізняльний поріг).
Сприймання звуків різної частоти. Людина сприймає звуки у діапазоні 20 -1 б 000 Гц (10 - 11 октав). Звуки
частотою до 20 Гц називають інфразву-ками, а понад 16 кГц — ультразвуками. Здатність людини і багатьох тварин розрізняти звуки за тонами є надзвичайно високою: поріг розрізняння частот оптимального діапазону (1 кГц) становить 0,3% (3 Гц).
Звук однієї частоти коливань називають тоном (чистим тоном). Більшість звуків створюється кількома частотами. Звичайно поєднуються основна частота і кілька гармонік — частоти, кратні основній за значенням, — це музичні звуки.
Звук, який складається з пепов'язапих між собою частот, називають шумом (білим шумом), якщо в ньому однаковою мірою представлені всі основні частоти в діапазоні чутності.
Чутливість людського вуха до звуків різної висоти неоднакова. Найкраще людина чує звуки частотою 2 — 5 кГц, хоча добре сприймає і звуки нижчої частоти, починаючи з 300 — 500 Гц. Звукові коливання саме цих частот людина використовує для своєї мови, і тому їх вона чує найкраще.
Сприймання сили (інтенсивності) звуку. Сила звуку — це кількість енергії, яка проходить крізь одиницю поверхні за одиницю часу (ват на метр квадратний). Сила звуку, що надходить до вуха, кодується частотою імпульсів у волокнах завиткового нерва, а також кількістю збуджених нейронів, оскільки слабкі звуки збуджують лише пайчутливіші нейрони, а сильні звуки залучають нервові волокна з вищим порогом подразнення.
Пряме визначення сили звуку є досить складною справою, тому на практиці замість сили використовують звуковий тиск як силу, що діє на площину, розміщену перпендикулярно до руху звукової хвилі. Мінімальний тиск, який здатна сприйняти людина, становить 2-Ю"5 Н/м2. Сила звуку під час звичайної розмови становить приблизно 10_1 Н/м2, а звук, що викликає больове відчуття у вухах, — ЗО Н/м2. Проте замість абсолютних значень звукового тиску звичайно застосовують таке поняття, як рівень звуко
вого тиску (О, який виражається у децибелах:
1=20- Іод (Р/Р0),
де Р — середнє значення звукового тиску, а Р0 — поріг чутності людини, тобто 2-Ю"5 Н/м2 (відносний нуль). Отже, зміна сили звуку у 10, 100, 1000 і 10 000 разів відповідно становить 10, 20, ЗО і 40 дБ.
Абсолютна слухова чутливість — це мінімальна сила звуку, який чує людина у 50% випадків його подачі. Поняття абсолютного слуху має інше значення. Його мають люди, які здатні точно пізнавати і позначати будь-який звук, навіть якщо немає звуку порівняння. Людське вухо розрізняє різницю в інтенсивності двох звуків однієї частоти силою 1 дБ.
Фізіологічна гучність — це суб'єктивно сприйнята сила звуку, яка виражається у фонах. Фон — це рівень звукового тиску топу, що порівнюється з еталонним топом частотою 1 кГц за однакової фізичної гучності їх звуку. Середній слуховий поріг здорової людини становить 4 фони.
Відчуття болю у вухах виникає за гучності звуку = 130 фонів. Звук такої сили може призвести до оборотної (чи необоротної) втрати слуху, оскільки при цьому ушкоджуються волоскові сенсорні еиітеліоцити спірального органа завитки або порушується мікроциркуляція в ній. У випадках тривалого впливу звуків інтенсивністю понад 90 дБ можлива звукова травма.
Визначення напрямку звуку. Людина легко визначає напрямок надходження звуку за допомогою бінаурального слуху (слухання обома вухами). В основі цього процесу лежить здатність слухової системи визначати різницю в силі звуків, що сприймаються обома вухами, якщо голова екранує звук, що надходить збоку. У випадках, коли довжина звукової хвилі дає змогу їй огинати головах — екран, напрямок звуку визначається) за запізненням надходження звуку .до другого вуха. Наша слухова система надійно визначає затримку звуку тривалістю
З • 10"5 с, що відповідає відхиленню джерела звуку від середньої лінії на 3° (за оптимальних умов можна розрізнити навіть кут 1,5°). Лише у випадках, коли звук надходить точно спереду або ззаду, людина повинна дещо повернути голову, щоб створити кут, необхідний для сприймання напрямку звуку. Якщо у навушниках скомпепсувати затримку звуку збільшенням сили сигналу, то людині видається, що джерело звуку знаходиться всередині голови.
У розрізнянні напрямку звуку бере участь також вушна раковина. Завдяки своїй формі вона сприяє чіткішому сприйманню напрямку звуку згори донизу і ззаду наперед. Людина з однобічною глухотою також здатна визначати напрямок звуку, але менш досконало і в будь-якому випадку повинна повертати голову.
При дослідженні центральних механізмів бінаурального слуху було виявлено у завиткових ядрах оливи довгастого мозку і нижніх горбках чотиригорбкової пластинки середнього мозку нейрони, які максимально збуджуються ири надходженні до обох вух звуку з різницею за часом або інтенсивністю. Кора великого мозку також бере участь в аналізі цих процесів: після руйнування слухової зони кори просторова звукова орієнтація порушується.
Ехолокація — це здатність організму орієнтуватись у просторі за допомогою звукових коливань. Ехолокація властива більшості хребетних тварин. Умовно можна виділити пасивну й активну ехолокацію.
Пасивна ехолокація полягає у визначенні за допомогою бінаурального слуху напрямку надходження звуку і його джерела. Це дуже ефективно і якісно роблять усі ссавці з розвиненими зовнішніми вухами. До пасивної ехолокації можна віднести і визначення перешкоди чи будь-якого об'єкта сприйманням відбитих від нього звукових хвиль, що виникли під час руху самого організму. Так, багато риб добре орієнтуються щодо розташування берега, різних предметів у воді, сприймаючи мехапорецеиторами бічної
Гравітаційна
сенсорна система
лінії відбиті інфразвукові коливання, створені рухом власного тіла. Незряча людина завдяки добре розвиненому слуху здатна визначати перешкоду за допомогою звуків, що виникли під час її крокування і відбились від перешкоди.
Активна ехолокація — це орієнтування у просторі відносно рухомих і нерухомих об'єктів за допомогою звуків, спеціально випромінюваних з цією метою організмом. Така форма ехолокації властива лише небагатьом видам вищих хребетних для орієнтування в просторі під час руху в темряві. Активну ехолокацію мають кажани, стрижі-саланган, птах гуаха-ро, деякі види китових. їхня здатність до активної ехолокації досягла такого високого розвитку, що стрижі, наприклад, легко визначають у темряві дротину діаметром менш як 1 мм, а комахоїдні кажани користуються своїм ехолокатором під час полювання па дрібних комах.
Стрижі та гуахаро, літаючи в темряві печер, здійснюють ехолокацію звуковими иосилами у вигляді коротких клацань звуковою частотою 4500 — 8000 Гц. Водночас кажани й дельфіни використовують для ехолокації ультразвуки частотою 50 — 80 кГц, хоча верхня межа ультразвукового діапазону, яку вони здатні сприймати, досягає 120-140 кГц.
Дослідження центральних механізмів ехолокації показало, що у кажанів аналіз ехосигиалів відбувається у передніх горбках чотиригорбкових пластинок покрівлі середнього мозку. Після їх зруйнування тварини втрачають здатність уникати перешкод, тоді як білатеральне видалення слухової зони кори у більшості кажанів не впливає на цю здатність.