

1. Стадія алергічної реакції, при якій виникає перший контакт з алергеномі розвивається сенсибілізація організму, називається:
А) імунологічна стадія,
Б) патохімічна стадія,
В) патофізіологічна стадія.
2. Стадія алергічної реакції, при якій відбувається синтез медіаторів алергічних реакцій, називається:
А) імунологічна стадія,
Б) патохімічна стадія,
В) патофізіологічна стадія.
Б. Ситуаційні задачі для самоконтролю:
1. Відомо, що перші ознаки ревматизму виникають через два тижні після перенесеної ангіни. Чому? До алергічних реакцій якого типу відноситься це захворювання?
2. Відомо, що при анафілактичному шоці ознаки хвороби розвиваються майже негайно після контакту з алергеном. Чому? До алергічних реакцій якого типу відноситься це захворювання?
VI. ЗМІСТ ТЕМИ ЛЕКЦІЇ В СТРУКТУРНО – ЛОГІЧНИХ СХЕМАХ.
VII. МАТЕРІАЛИ АКТИВІЗАЦІЇ
СТУДЕНТІВ.
Таблиці:
Стадії КПР.
Види ком
Мультимедійне забезпечення:
слайди,
відеоматеріал.
Роздатковий матеріал :
- набір таблиць з теми лекції,
- набір схем з теми лекції,
- тестові завдання,
- ситуаційні задачі.
VIІІ. МАТЕРІАЛИ ДЛЯ САМОПІДГОТОВКИ СТУДЕНТІВ.
ЛІТЕРАТУРА ПО ТЕМІ, ЩО ВИКЛАДЕНА В ЛЕКЦІЇ.
1. Навчальна:
А) Основна - І. Я. Федонюк „ Анатомія та фізіологія з патологією”,
Б) Допоміжна - В. І. Філімонов « Фізіологіялюдин»
Г. М. Чайченко « Фізіологіфя людини»
І. С. Кучеров «Фізіологія людини»
С. А. Георгієва « Фізіологія»
2. Методична:
- В. Є. Мілерян « Методичні основи підготовки та проведення навчальних занять в медичних ВУЗах»,
- Збірник нормативно - методичних матеріалів з організації навчального процесу у вищих медичних закладах освіти.
Оптичні середовища ока, або діоптричний апарат, — це складна система лінз, яка формує на сітківці обернене і зменшене зображення предметів зовнішнього світу. Ця система складається з прозорої рогівки, передньої і задньої камер, райдуж-ки, яка утворює зіницю, кришталика і склистого тіла, що займає більшу частину очного яблука. Склисте тіло — це прозорий гель, склиста волога і склиста строма — колоїдний розчин колагену й гіалуронової кислоти.
Фізіологія зору 41
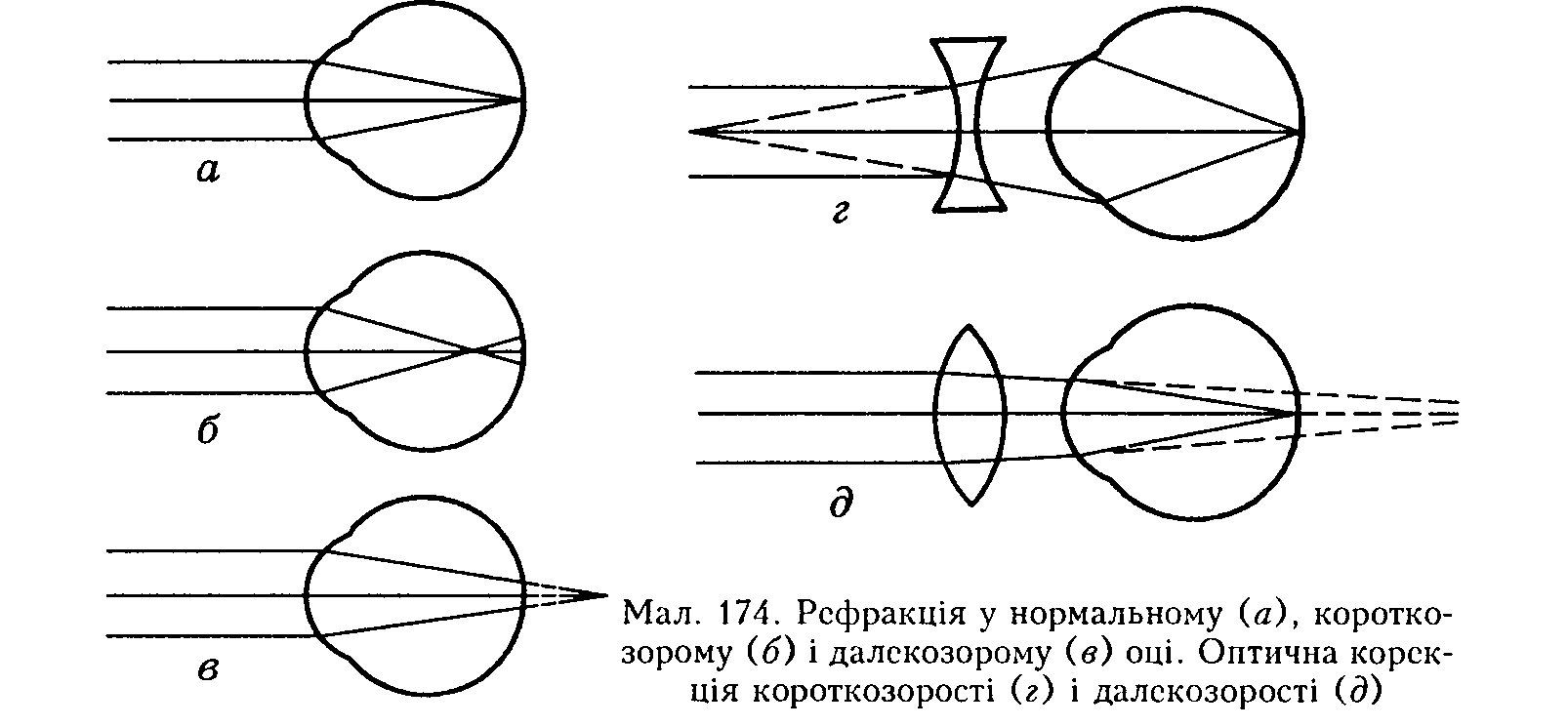
Для того щоб на сітківці виникло чітке зображення, світло, що падає па око, мусить заломитись так, щоб сфокусуватись на ній (мал. 174, а). Це заломлення відбувається переважно у рогівці й кришталику. В оці людини заломлення у рогівці приблизно у два рази більше, ніж у кришталику (загальна заломлювальна сила кришталика близько 19,1 дптр*). Заломлювальна сила всієї оптичної системи ока людини становить 59 дптр ири розгляданні далеких і 70,5 дптр — близько розташованих предметів.
Акомодація ока. Оскільки рогівка розміщена на фіксованій відстані від сітківки (24,4 мм) і не змінює своєї кривини й заломлювальної сили, то для фокусування зображення на сітківці різновіддалених предметів необхідна корекція заломлення створюється кришталиком. У риб вій майже сферичний, з високим коефіцієнтом заломлення і малою фокусною відстанню, тому що показник заломлення води майже такий, як і рогівки, і на ній заломлення не відбувається. Око риб у спокої акомодоване па коротку відстань, тому акомодація ока для них полягає у встановленні його для бачення вдалину. Це досягається тим, що кулеподібний кришталик спеціальним м'язом підтягується ближче до сітківки.
У земноводних форма кришталика також не змінюється при акомодації і спеціальний м'язовий апарат притягує його дещо вперед, встановлюючи око для дуже близького бачення (на відстань викидання язика), що необхідно для полювання на комах. У плазунів і птахів акомодація ока для близького бачення відбувається через зміну форми кришталика.
У ссавців цей процес також здійснюється пластичною зміною форми (кривини) кришталика. Форма кришталика ссавців залежить від його еластичності і впливу на його сумку. Коли війковий м'яз скорочується, він протидіє силам пружності, що діють на кришталик через волокна цинно-вих зв'язок, так що натягнення сумки кришталика зменшується. Внаслідок цього кривина передньої частини кришталика збільшується і його заломлювальна сила зростає, тобто кришталик виявляється в етапі акомодації па сприймання близьких об'єктів. Якщо ж війковий м'яз розслаблюється, то кривина кришталика і його заломлювальна сила зменшуються. У такому стані здорове око дає на сітківці чітке зображення далеких об'єктів.
Порушення рефракції ока. Якось Г. Гельмгольц пожартував, що коли б йому принесли оптичний прилад, сконструйова
35Ц ФІЗІОЛОГІЯ СЕНСОРНИХ СИСТЕМ
ний так незугарно, як око, то він би повернув його майстру назад. Проте фізіологічна "недосконалість" ока як фокусуючого оптичного пристрою значною мірою компенсується пейроииими механізмами контрастування. Які ж розрізняють оптичні "недосконалості" ока й аномалії його рефракції?
У людей найчастіше визначають такі дві аномалії рефракції ока: короткозорість (міопія) і далекозорість (гінермегронія) (див. мал. 174, б, в). Вони пов'язані з природженими відхиленнями від нормальної довжини очного яблука. В першому випадку поздовжня вісь ока перевищує 22,4 мм (відстань між полюсом рогівки і центральною ямкою нормального ока), тому зображення фокусується перед сітківкою (див. мал. 174, б), а на сітківці воно нечітке. У другому, навпаки, ири малій довжині очного яблука зображення проектуватиметься за оком (див. мал. 174, в), і людина також бачить погано. Крім того, з віком кришталик стає менш еластичним унаслідок певної дегідратації, його заломлювальна сила і діапазон акомодації зменшуються. Найближча точка нормального бачення (7 см) поступово відсувається, розвивається стареча далекозорість.
Астигматизм. Поверхня рогівки несиметрична відносно оптичної осі і звичайно кривина у вертикальній площині дещо більша, ніж у горизонтальній, що спричинює різницю заломлювальної сили променів, які йдуть під різними кутами. Якщо ця різниця не перевищує 0,5 дптр, то такий астигматизм називають фізіологічним (норма). Більша різниця зумовлює спотворене зображення на сітківці.
Аберація. Як у всіх простих лінз, у рогівки та кришталика фокусна віддаль неоднакова для різних їх частий — центральна частина, крізь яку проходить оптична вісь, має найбільшу фокусну віддаль. За рахунок цього виникає сферична аберація, що призводить до нечіткого зображення. Проте цей недолік компенсується розмірами зіниці: чим менший розмір зіниці, тим чіткішим буде зображення, оскільки в цьому разі периферичні частини оптичної системи майже не беруть участі у формуванні зображення. Так в оці коригуються недоліки, зумовлені сферичною аберацією.
Як усі прості лінзи, діоптричний апарат ока заломлює світло з коротшою довжиною хвилі більше, ніж з довшою. Це явище називають хроматичною аберацією. Тому для точного фокусування червоних частин предметів потрібен більший ступінь акомодації, ніж для синіх. Саме тому сині предмети видаються нам більш віддаленими, ніж червоні, навіть якщо вони розташовані на одній площині і відстані. Архітектори готичних церков часто використовували цю фізіологічну ілюзію при створенні кольорових вітражів: фон вони роблять синього кольору, а фігури іншими барвами, і тому видається, що ці фігури ніби виступають з фону.
У людей похилого віку вміст води у кришталику знижується, і він може втрачати свою прозорість (стареча катаракта). Якщо таким людям видалити кришталик, то вони можуть нормально бачити лише в окулярах з лінзами 13 дптр.
Коригування порушень рефракції ока. Для коригування коротко- і далекозорості застосовують окуляри відповідно з увігнутими й опуклими лінзами (див. мал. 174, г — д). Астигматизм коригують залежно від його форми.
Реакції зіниці. Середня яскравість навколишнього середовища коливається у широких межах. Зорова система пристосовується до таких коливань освітленості за допомогою реакцій зіниці. У людини зіниці обох очей мають зазвичай круглу форму і однаковий розмір, який з віком зменшується. При постійному зовнішньому освітленні кількість світла, що потрапляє до ока, є пропорційною площі зіниці. При зниженні інтенсивності зовнішнього освітлення зіниці рефлекторно розширюються. У молодих людей діаметр зіниці може змінюватись від 1,5 до 8 мм, що зумовлює зміну освітлення сітківки приблизно у ЗО разів. До того ж при звуженні зіниці зростає глибина різкості, як у фотоапараті ири зменшенні отвору діафрагми.
Якщо ири денному освітленні людина заплющує очі на 10 —20 с, то діаметр зіниці зростає. Якщо освітлювати лише одне око,
його зіниця звужується (пряма реакція на світло), але при цьому звужується й зіниця иеосвітлепого ока (співдружня реакція на світло). Реакція зіниць на світло є одним з механізмів адаптації, що забезпечує оптимальне освітлення фоторецепторів сітківки.
У пічних тварин зіниця у напівтемряві спалахує зеленкуватим блиском, оскільки в них добре розвинений шар пігментного епітелію сітківки, що відбиває світло, яке проходить крізь зіницю. Таким чином світло двічі проходить крізь зорову частину сітківки, що має велике значення для пічних тварин. Цей шар розміщений у верхній половині сітківки, тому на нього потрапляють головним чином промені від поверхні землі, що дає змогу тварині бачити в темряві навколишні "земні" предмети.
Сльозова рідина. Надзвичайно чутлива і ніжна прозора рогівка постійно контактує з повітрям навколишнього середовища, багатога па часточки пилу й різні шкідливі речовини. Щоб захистити її від їх ушкоджувального впливу, зовнішня поверхня рогівки вкрита тонким шаром сльозової рідини. Крім того, завдяки сльозовій рідині поліпшуються оптичні властивості рогівки.
Сльозова рідина у невеликих кількостях постійно виробляється сльозовими залозами. Завдяки рухам повік вона рівномірно розподіляється па рогівці й кон'юнктиві. Частина цієї рідини випаровується, а решта спливає сльозовими протоками до сльозового озера, а потім до порожнини носа. Сльозова рідина захищає рогівку й кон'юнктиву від висихання і одночасно є своєрідним мастилом між очним яблуком і повіками.
На смак сльозова рідина солона, за складом близька до плазми крові. Сльозова рідина містить також ферменти, які мають бактерицидну дію, захищаючи око від інфекції. У людини секреція сльозової рідини виконує ще одну важливу функцію — бере участь у вираженні емоцій, полегшує стресовий стан.
Більшість рефлексів, що зумовлюють збільшення секреції сльозової залози, за
Фізіологія
зору
пускається з рецепторів, розміщених в очній ділянці, інформація від яких через трійчастий нерв передається до стовбура мозку. Проте підвищену секрецію сльозової залози можуть спричинити й сторонні стимули, наприклад зубний біль, механічне подразнення слизової оболонки носа, різкі смакові стимули або пахощі, а також чинники, що викликають кашель. Секрецію сльозової рідини регулює автономна нервова система.
12.2.3. СВПТЮСПРИЙМАЛЬНА СИСТЕМА ОКА
Основним елементом світлоснриймаль-ної системи ока є сітківка — внутрішня (сенсорна) оболонка очного яблука, яка в онтогенезі розвивається з випинання частини проміжного мозку за межі черепа. Вона знаходиться па внутрішній поверхні задньої частини очного яблука і безпосередньо прилягає до його судинної оболонки.
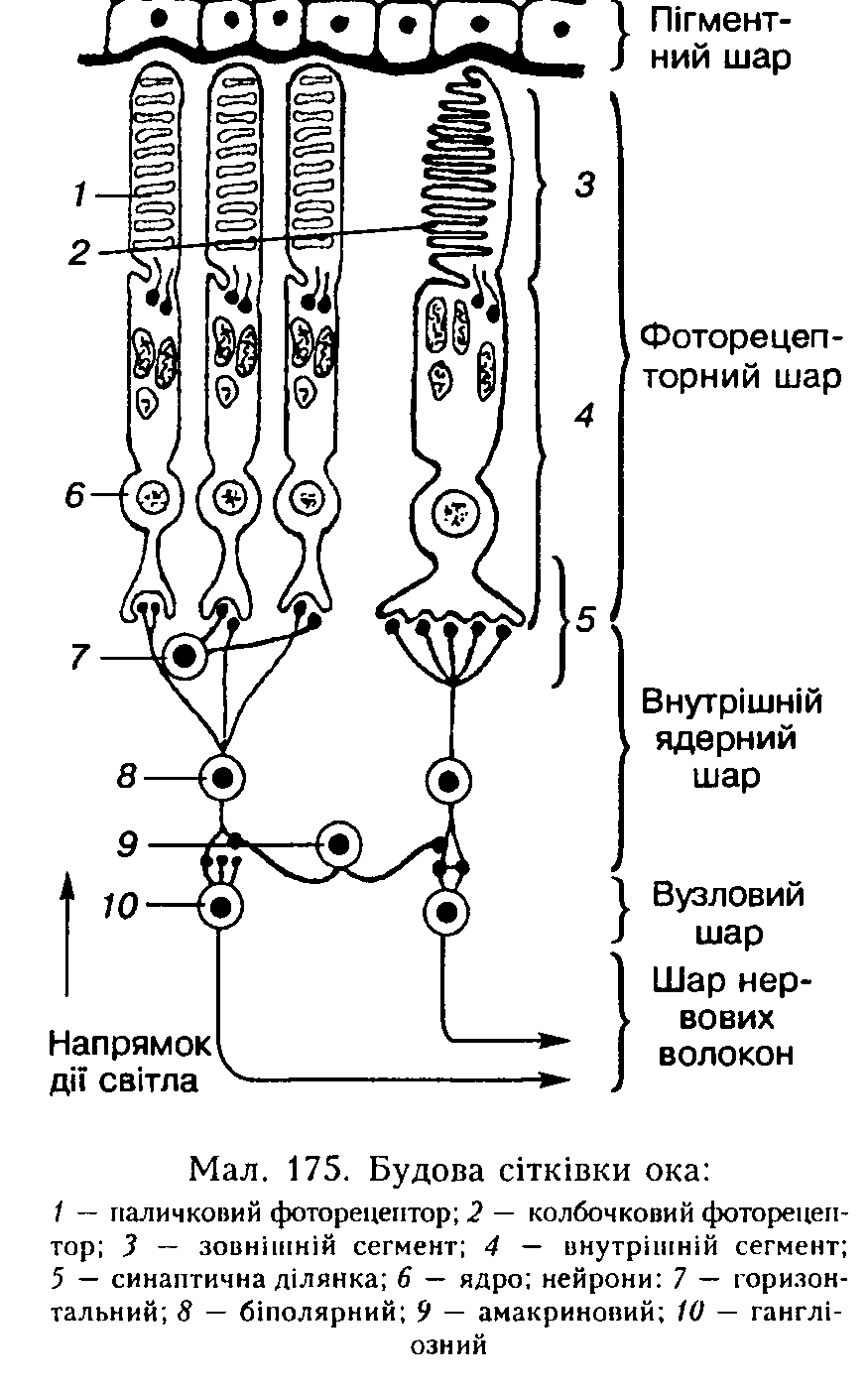
Будова сітківки. Сітківка (мал. 175) складається зі сліпої і зорової частин. Зорова частина сітківки складається з двох шарів: пігментного і нервового. До складу нервового шару входить фотосенсорпий шар, що складається з паличкових і кол-бочкових фоторецепторів, два шари нейронів і шар нервових волокон. У трьох останніх шарах знаходяться також гліаль-11 і клітини. У людини налічують 130 мли фотосенсорпих клітин (з них 6 — 7 мли кол-бочкових), від яких сигнали надходять майже до 1 мли гані ліозпих нейронів сітківки, тобто в середньому кожний гап-гліозпий нейрон отримує інформацію від 120 паличкових і б колбочкових фоторецепторів.
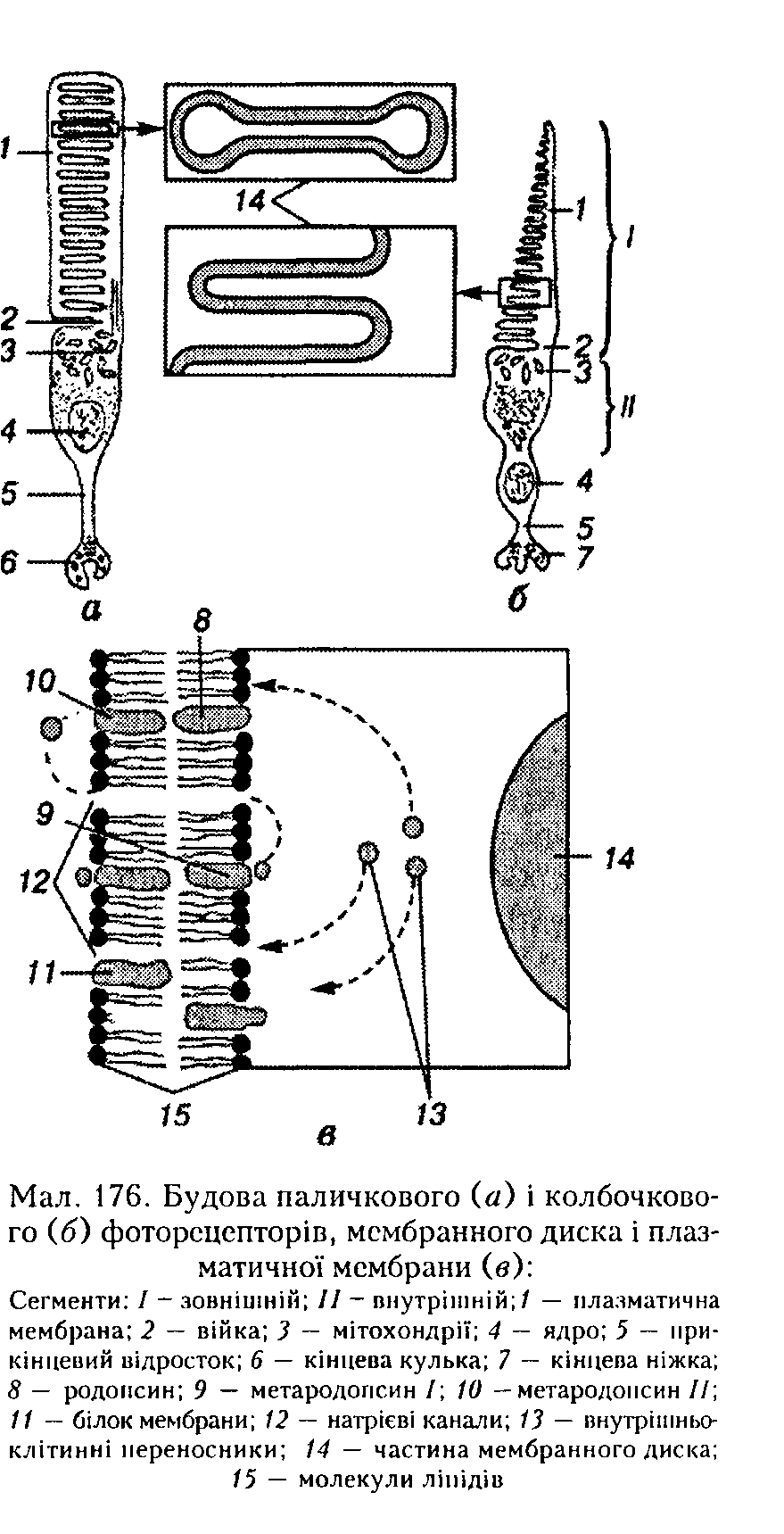
Фотосенсорпий шар є складовою нервового шару зорової частини сітківки. Він складається з паличкових і колбочкових фоторецепторів. Вони побудовані із зовнішнього і внутрішнього сегментів, з'єднаних сполучною ніжкою (війкою), ядер-иоплазматичиого тіла, нретерміиальиого відростка і складного синаитичиого закінчення — кінцевої кульки паличкового і
кінцевої ніжки колбочкового рецепторів (мал. 175, 176). Зовнішній сегмент паличкового фоторецептора— це вузька трубка, заповнена численними тонкими мембранними дисками, замкненими з усіх боків плоскими мембранними мішечками, відокремленими від плазматичної мембрани і оточеними нею. І трубка, і диски утворені двошаровою мембраною того самого типу. Диски містять більшість мембранних білкових молекул, які беруть участь у поглинанні світла і генерації рецепторного потенціалу.
Будова колбочкових фоторецепторів практично така сама, як і паличкових, хоча їхній зовнішній сегмент за діаметром менший від внутрішнього і звичайно має конічну форму, а внутрішній сегмент коротший і товщий, ніж у паличкових. Крім того, мембранні диски зовнішнього сегмента колбочкових фоторецепторів є складками зовнішньої частини плазматичної мембрани і міждисковим простором, сполученим з позаклітинним середовищем.
Нейронні шари сітківки. Паличкові і колбочкові фоторецептори утворюють сииаитичні сполучення з біполярними нейронами внутрішнього ядерного шару сітківки, короткий аксон яких закінчується синансом на гапгліозних (мультииолярних) нейронах вузлового (гаигліозного) шару сітківки. Зазвичай кілька біполярних ией
Фізіологія
зору
ропів конвергують па одному гапгліозному нейроні, утворюючи збудливу зону.
У сітківці є також інші типи нейронів, які не входять до цього трииейропио-го ланцюга зорового аналізатора. Це горизонтальні та амакринові нейрони (див. мал. 175). Горизонтальні нейрони (внутрішній ядерний шар) не мають типового аксона, а будова їхніх синапсів така, що не можна встановити напрямок передачі сигналів. Тому вважають, що горизонтальні нейрони отримують вхідні сигнали від паличкових фоторецепторів, а вихідний сигнал спрямовується або назад до них, або до біполярних нейронів, або до тих й інших.
Амакринові нейрони розміщуються між біполярними. Вони мають різноманітну форму і виділяють близько 20 медіаторів. У них також немає аксонів, проте їхні дендрити здатні утворювати пресииаитич-иі закінчення па інших клітинах. Так, контакт біполярних нейронів з гангліозними здійснюється за допомогою амакринових нейронів, які утворюють обернений зв'язок з біполярними і відіграють важливу роль у передачі сигналу до гангліозиих нейронів (див. мал. 175).
Вихідний нейрониий шар сітківки — це шар гангліозиих мультиполярних нейронів, аксони яких проходять крізь сітківку і білкову оболонку ока — склеру і утворюють зоровий нерв. Волокна присередпьої частини зорового нерва перехрещуються, і після перехрестя у складі зорового шляху усі волокна прямують до трьох підкіркових центрів зору: бічного колінчастого тіла, подушки таламуса і сірого шару верхнього горбка покрівлі середнього мозку. У місці проходження зорового нерва крізь сітківку фоторецепторів немає — це диск зорового нерва, або сліпа частина сітківки (сліпа пляма). У центрі сітківки ( по оптичній осі ока) розміщена центральна ямка (жовта пляма). Тут зосереджені лише кол-бочкові фоторецептори, і кожен зв'язаний тільки з одним гапгліозиим нейроном (див. мал. 175). Ось чому гострота зору в зоні центральної ямки є максимальною. Коли об'єкт фіксується оком, його зображення потрапляє саме па центральну ямку.
Сітківка в оці хребетних тварин розміщена таким чином, що її пігментний шар прилягає до судинної оболонки і внутрішньої поверхні склери, а гаигліозпий шар контактує зі склистим тілом. При цьому світло, щоб потрапити до фотосенсорио-го шару, повинно пройти всі шари сітківки і відбитись від пігментного шару. Таке око називають інвертованим, на відміну від неіпвертоваиого ока у головоногих молюсків.
Фотохімічні процеси в фоторецепторах сітківки. Зовнішній сегмент фоторецептора містить зорові пігменти — основну частину молекулярного механізму, що сприймає світло і запускає потік інформації до нейронів сітківки. Внутрішній сегмент генерує енергію і поновлює молекули зорового пігменту, необхідні для зовнішнього сегмента. Крім того, внутрішній сегмент формує синаптичні закінчення для зв'язку з іншими нейронами сітківки.
Фотосенсорпий білок паличкових фоторецепторів — це родопсин, зв'язаний з мембраною дисків. Вій має пурпуровий колір (відбиває сині й зелені промені), його називають зоровим пурпуром. Родопсин є однією з найбільш інтенсивно забарвлених сполук органічного світу, яка має дуже широкий діапазон поглинання. Вій активно поглинає кванти світла у широкій смузі частот видимого спектра і започатковує цілий ланцюг хімічних реакцій, які в кінцевому підсумку забезпечують зір. Розглянемо в скороченому вигляді цей каскад хімічних реакцій.
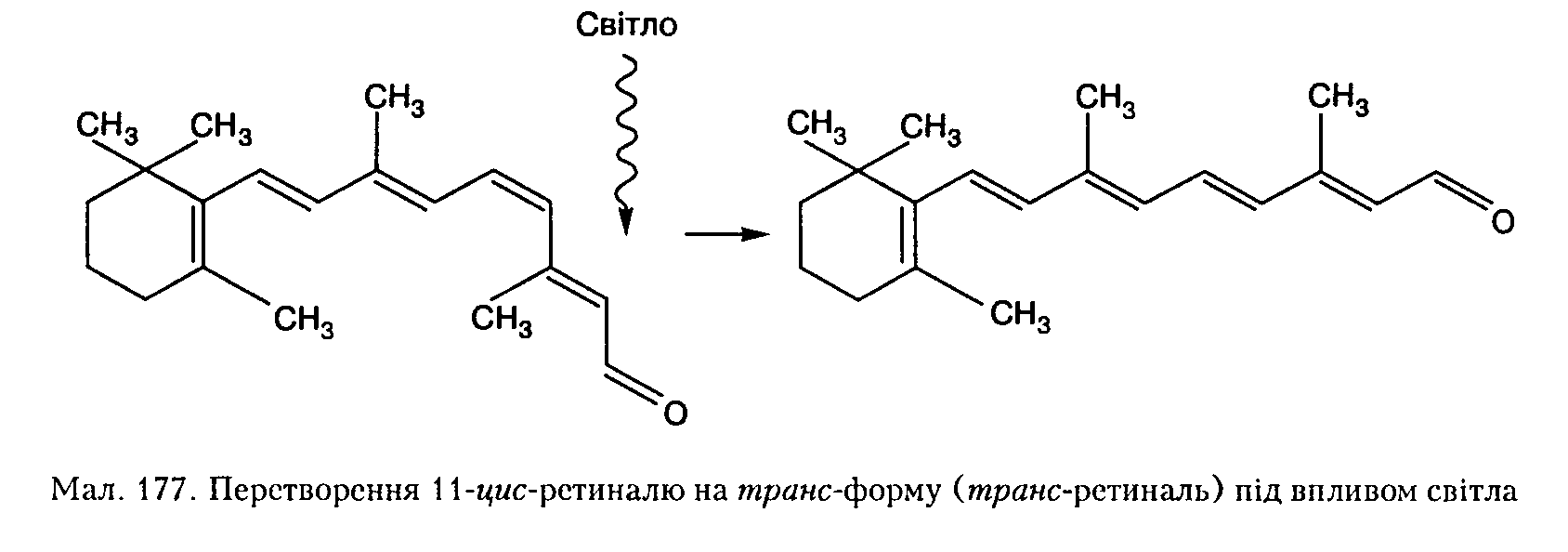
Родопсин складається з двох компонентів — хромофора (11-цис-ретиналь), який визначає інтенсивність поглинання світла, і безбарвного білка опсину (ско-тоисипу). Хромофор є похідним ретинолу (віт. А), а опсии є ферментативним білком — каталізатором хромофора. Поглинання фотона світла хромофором активує опсип і запускає каскад біохімічних реакцій.
Насамперед внаслідок цис-транс-ізомерн-зації 11-цг/с-ретиналь набуває транс-фор-ми (мал. 177). Поглинувши квант світла, 11-цг/с-ретиналь набуває електронно збудженого стану. Час ізомеризації 11-цг/с-рети-
налю становить 10"12 с. Ця иікосекупдна ізомеризація можлива безпосередньо лише у білковій макромолекулі, у розчині родопсину вона здійснюється у 1000 разів повільніше.
Білок сприяє ретиналю швидко переходити не тільки з цис- до трянс-форми, а й навпаки — від транс- до г^г/с-форми. Цей зворотний процес називають фоторегене-рацією, він є типовим для фоторецепторів безхребетних тварин. У хребетних тварин основний шлях регенерації родопсину — "темповий": етгрянс-ретииаль відщеплюється від білка опсииу і прямує до клітин пігментного шару сітківки, а на його місце з них надходить повий 11-^г/с-ретиналь. Лише цей ізомер здатний зв'язуватися з білком і утворювати оновлену молекулу родопсину.
За сучасними уявленнями, послідовність перетворення родопсину під виливом світла є такою (в дужках діапазон поглинання): під час перетворення метародопсипу І на метародопсин II. Саме па цій стадії молекула родопсину, яка поглинула квант світла, взаємодіє з білками мембрани паличкового фоторецептора, активуючи їх. Це призводить до збільшення проникності мембрани для йонів Са2+. Останні шляхом дифузії або за допомогою молекул-нере-носииків (мал. 176, в) досягають натрієвих каналів у мембрані зовнішніх сегментів паличкових рецепторів і спричинюють замикання цих каналів, тобто зменшують провідність цієї мембрани для №+, що зумовлює її гіперполяризацію.
Отже, копформаційиі зміни молекули родопсину зумовлюють генерацію на мембрані зовнішнього сегмента паличкового фоторецептора гіперполяризаційного РП. Амплітуда цього потенціалу звичайно пропорційна інтенсивності світлового стимулу. Таким чином, па відміну від інших органів чуття у паличкових фоторецепторах під впливом адекватного зовнішнього подразника виникають гінериоляризаційпі зрушення.
Який же механізм генерації цього потенціалу? Порівняно з іншими нервовими чи рецепторними клітинами мембрана паличкового фоторецептора у темряві має досить високу натрієву провідність, що зумовлює появу темпового струму. Цей струм у зовнішньому сегменті паличкового фоторецептора забезпечується молекулами цАМФ, які підтримують відкритими йоииі канали мембрани. Світло активує фосфодіестеразу, що викликає зниження концентрації цАМФ і закриття каналів.

Найбільша коиформаційиа перебудова білкової частини родопсину відбувається
Фізіологія
зору
Темповий струм зменшується, відбувається гіперполяризація, яка додається до розглянутої вище кальцієвої гіперполяри-зації.
Гіперполяризаційпий РП, що виникає під впливом світла, є унікальним явищем, для розуміння якого потрібно враховувати, що нормальним режимом роботи паличкового фоторецептора є підрахунок окремих квантів світла (фотонів). У відповідь на поглинання фотона паличковий фоторецептор генерує тільки короткий поодинокий електричний імпульс. Проте оскільки світло має квантову природу, тобто стимул фактично є перервним (дискретним), то паличковий рецептор використовує звичайний для нервової системи імпульсний код, реагуючи на кожний фотон квантовим сплеском.
Оскільки поглинання фотона не супроводжується появою ПД, то імпульсний сигнал, що виникає, поширюється через аксон електротоиічио (з декрементом), але завдяки дуже коротким аксонам клітин у межах сітківки затухання квантового сплеску є незначним. Це забезпечує достатню амплітуду сплеску, який викликає одноразове спрацьовування синапсу. Отже, квантова природа світла дала можливість відмовитися від додаткової надбудови — ПД і обмежитися лише квантовим сплеском, який і поширюється електротоиічио до синапсу паличкового фоторецептора.
Після припинення освітлення ока у темряві відбувається ресинтез родопсину, для чого потрібно, щоб до сітківки потрапив цис-ізомер ретинолу (віт. А), з якого утворюється ретииаль. Тому за браком в організмі ретинолу (а також йопів цинку) розвивається захворювання гемералопія (куряча сліпота).
У колбочкових фоторецепторах виявлено світлочутливий пігмент йодопсин, який має два максимуми поглинання — 562 і 370 пм. Структура йодопсипу дуже нагадує родопсин: йодопсин складається з 11-цг/с-ретиналю і білка фотопсину, що відрізняється від скотонсииу паличкових фоторецепторів.
Відповідно до зорових пігментів різняться й функції обох видів фоторецепторів. Оскільки паличкові фоторецептори з їх родопсином набагато чутливіші до світла, ніж колбочкові, то вони функціонують при слабкому денному і нічному освітленні (присмерковий зір), а колбочкові — при звичайному денному освітленні. У сутінках і при світлі зірок предмети здаються безбарвними, відрізняючись лише яскравістю (скотопічний зір), а вдень при яскравому освітленні око розрізняє не стільки яскравість, скільки переважно колір (фото-пічний зір). Крива спектральної чутливості ока під час переходу від скотоиічпого до фотопічного зору зміщується до коротших хвиль.
Поширення сигналу в сітківці. Під
виливом фотонів світла в колбочкових і паличкових фоторецепторах виникає рецепторний потенціал у вигляді їх гі-нерноляризації, амплітуда якої залежить від інтенсивності світла, що надає на фоторецептор, і яка в цьому разі є ознакою активного стану рецептора, а не його гальмування. Цей сигнал електротоиічио передається на біполярний нейрон, який також гінерноляризується (мал. 178). Переходячи від біполярного до гангліоз-иого нейрона, збудження трансформується з аналогової форми (гіперполяризаційпий РП) на імпульсну (ПД). Передача сигналу від біполярних нейронів до гангліозиих здійснюється також через амакринові нейрони (див. мал. 175). Вони є типовими нейронами, які у відповідь на гіперполяризаційпий стимул від біполярних нейронів генерують градуальну деполяризацію з поодинокими ПД на її верхівці. А вже гаигліозні нейрони на тлі їхньої деполяризації продукують пачки ПД, кількість яких у пачці пропорційна інтенсивності освітлення. Ця інформація у вигляді імпульсної активності через зоровий нерв надходить до ЦНС.
Горизонтальні нейрони також відіграють певну роль в обробці зорової інформації. Вони здійснюють просторову сумацію сигналів від фоторецепторів у межах рецептивного поля. Деякі з них (нейрони Ь) завжди відповідають гіпериоляризацією на дію світла будь-якої довжини хвилі, а інші
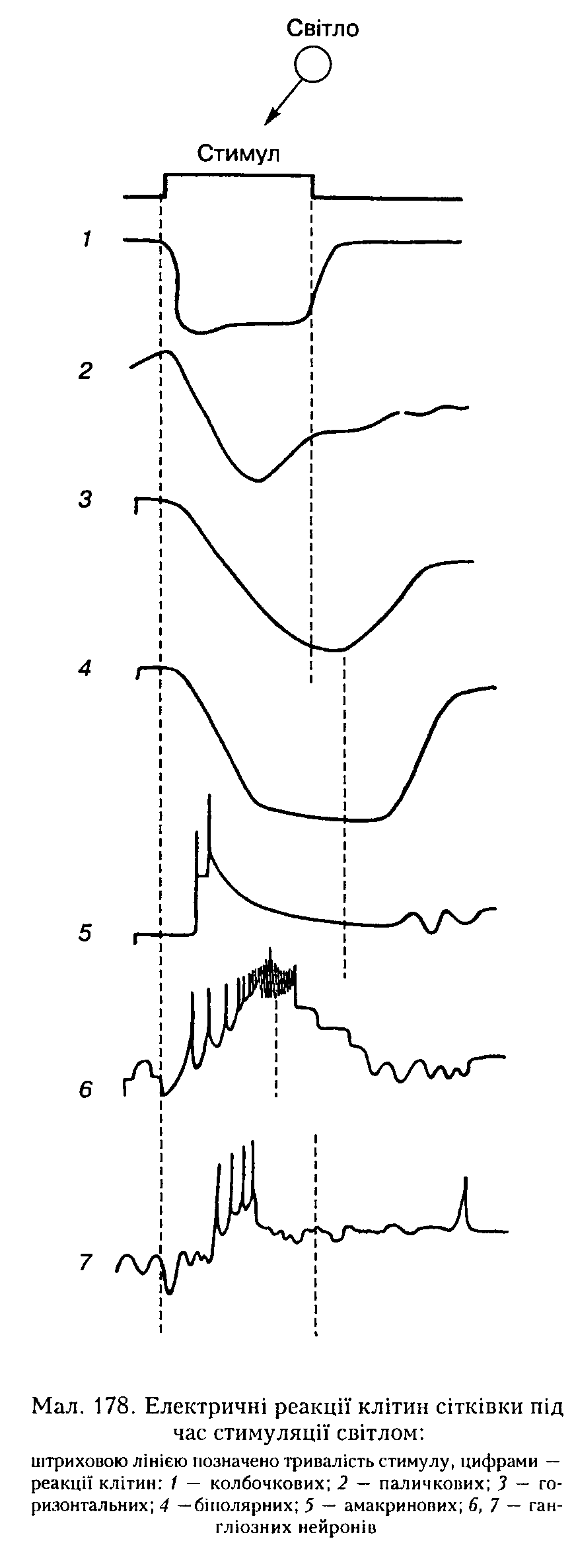
(нейрони С) реагують гінер- або деполяризацією залежно від довжини хвилі подразнювального світла.
Рецептивні поля гінгліозних нейронів. Функцію гангліозиих нейронів вузлового (гапгліозного) шару сітківки вивчають, реєструючи електричну активність окремих волокон зорового нерва. На кожному такому нейроні коивергує величезна кількість інших нейронів сітківки. Шляхом посилання дуже вузького пучка світла па різні точки сітківки можна знайти ту її ділянку, подразнення якої світлом спричинюватиме збудження або гальмування певного гіигліозиого нейрона. Ця ділянка і є його рецептивним полем. По-перше, гаигліозні нейрони реагують па освітлення сітківки короткочасно: лише на вмикання світла (нейрони-оп), лише па вимикання (нейрони-оЦ) і па вмикання й вимикання світла (нейрони-оп-о({). По-друге, нейрони рецептивного поля, розміщені в його центрі й на периферії, реагують на освітлення протилежним чином: якщо у відповідь па освітлення центра рецептивного поля иейрон-оя збуджується, то під час дії світла па периферичні частини цього поля цей нейрон гальмується. Відповідно він гальмується па вимикання світла в центрі рецептивного поля і збуджується па його периферії. Описане явище дістало назву бічного гальмування, воно зумовлене реципрокними взаємозв'язками між центральними і периферичними нейронами рецептивного поля.
Електроретинограма. У 1859 р. Е. Дюбуа-Реймон встановив, що між передньою і задньою частинами ока існує різниця потенціалів, яка змінюється під час освітлення. Запис цієї сумарної відповіді сітківки па освітлення називається електроретинограмою (ЕРГ) і складається з кількох коливань (хвиль) потенціалу (мал. 179), які відображають різні процеси: а-хвиля — початкове негативне коливання, зумовлене сумацією РП* фоторецепторів і горизонтальних нейронів; велика позитивна б-хвиля, зумовлена активізацією гліальних клітин сітківки; тривала позитивна в-хви-ля відображає зміни МП пігментоцитів —
Фізіологія
зору
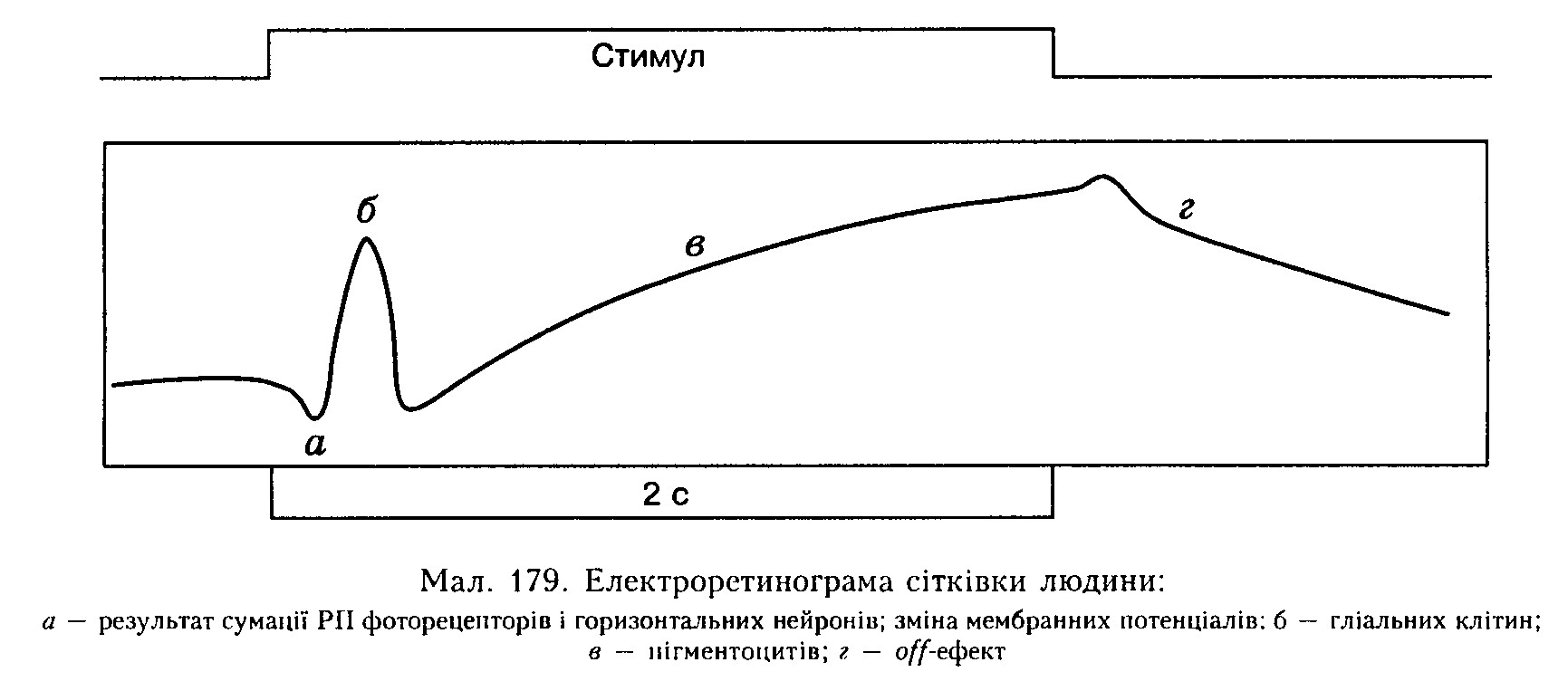
клітин пігментного шару сітківки на вмикання світла і г-хвиля — на вимикання світла, або так званий о/'/'-ефект. Та обставина, що ЕРГ відображає активність більшості елементів сітківки, дала змогу використовувати цей показник в офтальмології для діагностування деяких очних хвороб.
Світлова і темнова адаптація. Якщо загальне освітлення змінюється, то зорова система пристосовується до нових умов, змінюючи свою чутливість. Коли людина виходить з яскраво освітленої кімнати на темну вулицю, то в перший момент вона не може розрізняти навколишні предмети. Проте згодом їх контури стають помітними. Під час цієї темпової адаптації абсолютна чутливість зорової системи повільно зростає, і максимальної чутливості вона досягає лише через дві години перебування у повній темряві. Абсолютна чутливість у цьому разі становить (у розрахунку на один рецептор) 1—4 фотони світла за 1 хв.
Протилежний процес називають світловою адаптацією. Вона відбувається значно швидше, протягом кількох секунд (хоча на короткий час людина може бути повністю засліплена).
Вирішальну роль у процесах адаптації відіграють нейроииі механізми, які перемикають колбочковий зір на паличковий і навпаки. Крім того, важливим механізмом темпової і світлової адаптацій є зміна діаметра зіниці, що відбувається під час зміни рівня освітленості ока.
Й. В. Гете, який був не лише видатним поетом, а й природознавцем, описав таке явище: "Я повернувся до готелю повечеряти, і до моєї кімнати увійшла й стала на певній відстані від мене пишна покоївка з слінучо-білим обличчям, чорним волоссям і в червоній сукні. Я уважно придивився до неї. Після того як вона вийшла, я побачив на білій стіні проти себе чорне обличчя, оточене світлим пасмом, а одяг цієї цілком нової фігури видавався чудового синьо-зеленого кольору".
Це явище пояснюється локальною адаптацією і виникненням послідовних образів. Локальна адаптація виникає тоді, коли за сталої середньої освітленості обмежена ділянка сітківки має освітленість, яка відрізняється від середньої. Так, якщо протягом ЗО с фіксувати зір на якомусь зображенні, потім перевести погляд па рівний темний чи світлий фон, то протягом кількох секунд можна бачити негативний послідовний образ. У цьому послідовному образі те, що було па зображенні темним, видаватиметься світлим і навпаки. Чому так відбувається?
Ті ділянки сітківки, на які припадають темні місця зображення, за час фіксації зору стають чутливішими, ніж сусідні ділянки, на які припадали світлі місця зображення. Такі послідовні образи зберігаються досить тривалий час, якщо застосу
вати сильне освітлення сітківки або якщо "засвічування" (не дуже сильне) триває довго. У разі локальної адаптації до колірних стимулів утворюються послідовні образи, забарвлені у додаткові кольори.
У 1825 р. Ян Пуркіньє помітив, що вдень видаються яскравішими червоні кольори, а в сутінках — сипі. Це зрушення Пуркіньє пояснюється переходом від фото-нічного до скотопічпого зору під час темпової адаптації.