

Розпізнавання висоти тону
Основними характеристиками звукової хвилі є: частота, амплітуда і тривалість коливань. Сила звуку корелює з амплітудою звукової хвилі, а висота тону - з її частотою (кількість коливань за одиницю часу). Чим більша амплітуда, тим голосніший звук, чим вища частота коливань, тим вищий тон.
Вухо людини здатне сприймати звук за умови коливання повітря в діапазоні від 16 до 20000 Гц. Однак найбільша чутливість лежить у межах від 1000 до 4000 Гц - це діапазон людського голосу. У межах зони слухового сприйняття людина може відчувати близько 300 000 різних за силою і висотою звуків.
Припускають, що існує два механізми розпізнавання висоти тонів. У діапазоні частот понад 1000 Гц використовується механізм просторового
кодування. Звукова хвиля, яка являє собою коливання молекул повітря, поширюється у вигляді поздовжньої хвилі тиску. Ця рухома хвиля передається на пери- та ендолімфу; між місцем її виникнення та загасання існує ділянка, де амплітуда коливань є максимальною. Місцезнаходження цього амплітудного максимуму залежить від частоти коливань: за умови високих частот воно ближче до овальної мембрани, а в разі низьких частот - ближче до гелікотреми. У результаті цього амплітудний максимум для кожної частоти, яку людина чує, розташований у специфічній точці ендолімфатич-ного каналу. Так, амплітудний максимум для частоти коливань 4000 Гц знаходиться на відстані 10 мм від овального отвору, а 1000 Гц - 23 мм. Волокна основної мембрани, які розташовані на відповідній ділянці, і сенсорні клітини, що лежать на мембрані, коливаються найсильніше, так що нервовими волокнами, які відходять від них, іде найчастіша імпульсація. Цьому ж сприяє різна щільність основної мембрани уздовж завитки: вона є найбільшою біля основи. Градієнт щільності мембрани забезпечує поширення коливальних хвиль від основи до вершини. Початкова, найщільніша частина мембрани служить високочастотним фільтром - хвильові коливання далі не поширюються. За невеликої частоти коливань (до 1000 Гц) може використовуватися і телефонний (залповий) механізм кодування: у слуховому нерві виникає потенціал дії, частота якого відповідає частоті звукових коливань. Однак лабільність слухового нерва не дозволяє проводити коливання з частотою понад 1000 Гц, тому високі тони диференціюються тільки за допомогою просторового кодування.
Розпізнавання сили звуку
Сила звуку кодується кількістю збуджених нейронів і частотою імпульсів. Діапазон амплітуди коливань ендолімфи пов'язаний з амплітудою коливання мембран. З ростом амплітуди збільшується число збуджених рецептивних клітин. У разі слабких звуків у збудження втягується невелика кількість найчутливіших рецепторних клітин, а за умови посилення звуку до клітин, що перебувають на амплітудному максимумі, приєднуються клітини з вищим порогом збудження. Крім того, поріг збудження внутрішніх волоскових клітин більший, ніж зовнішніх. Тому в залежності від сили звукового подразнення змінюється співвідношення кількості збуджених внутрішніх і зовнішніх волоскових клітин.
Обробка звукової інформації у центральній нервовій системі
Збудження від рецепторних клітин органа Корті синаптично передається на дендрити нейронів спірального ганглію, розташованого в товщі спіральної пластинки. Аксони цих нейронів утворюють завиткову (кохлеар-ну) частину присінково-завиткового нерва (VIII пара черепних нервів). Волокна слухового (кохлеарного) нерва досягають вентрального і дорсального кохлеарних (завиткових) ядер довгастого мозку. Від вентрального ядра вони
направляються як до іпсі-, так і контрлатерального оливарнш комплексів. Дорсальний кохлеарний тракт переходить на протилежний бік і закінчується у ядрі латеральної петлі. Нейрони, які виходять з оливи і містять шляхи уже від обох завитків, також віддають колатералі в ядра латеральної петлі. Далі волокна спрямовуються до нижніх горбків чотиригорбкового тіла і до медіальних колінчастих тіл, які є головним релейним відділом слухової системи таламуса. Звідси збудження надходить до первинної слухової кори великого мозку, яка розташована у скроневій ділянці. Поруч з нею знаходяться нейрони, які належать до вторинної слухової зони кори.
Інформація, що міститься у звуковому стимулі, пройшовши всі зазначені ядра переключення, багаторазово (принаймні, не менше ніж 5-6 разів) "переписується" у вигляді нейронного збудження. На кожному етапі відбувається відповідний аналіз інформації, до якого нерідко підключаються сенсорні сигнали інших "неслухових" відділів ЦНС. У результаті можуть виникати рефлекторні відповіді, характерні для відповідного відділу ЦНС. Але тільки на рівні кори великого мозку інформація розпізнається як відповідний звук.
За звичайних умов чисті тони майже не зустрічаються. Нейрони вентрального ядра ще сприймають чисті тони, тобто збудження у них виникає під час дії чітко визначених тонів. У дорсальному ж ядрі лише невелика частина нейронів збуджується чистими тонами. Інші реагують на складніший стимул, наприклад, на змінні частоти, на припинення звуку тощо. На вищих рівнях поступово збільшується кількість окремих нейронів, що специфічно реагують на складні звукові модуляції. Так, одні нейрони збуджуються лише в разі змінної амплітуди звуку, інші - змінної частоти, треті - в разі зміни відстані до джерела звуку або його переміщення. Таким чином, щоразу під час дії складних звуків, що реально існують у природі, в нервових центрах виникає своєрідна мозаїка нейронів, які одночасно збуджуються. Відбувається запам'ятовування цієї мозаїчної карти, пов'язаної з надходженням відповідного звуку. Наповніше і досконало ці процеси відбуваються лише в кіркових відділах.
Від скроневої слухової ділянки кори (переважно від первинної) відходять низхідні шляхи практично до всіх підкіркових слухових ядер. До різних рівнів слухового тракту підходять шляхи і від неслухових відділів головного мозку. Так, до медіального колінчастого тіла йдуть шляхи від зорової зони кори та ядер мозочка. Широкі двобічні зв'язки слухових зон ЦНС, з одного боку, служать для поліпшення обробки слухової інформації, а з іншого - для взаємодії сенсорних систем.
Слухова орієнтація у просторі
Достатньо точна слухова орієнтація у просторі можлива лише завдяки існуванню бінаурального слуху (слуху двома вухами). Тут велике значення має те, що одне вухо знаходиться дальше від джерела звуку, ніж інше. Тому в обидва вуха звук надходить у різний час і з різною інтенсивністю,
що аналізується центральними відділами слухової сенсорної системи. Крім того, вушні раковини завдяки своїй формі сприяють концентруванню звуків та обмежують потік звукових сигналів з тильного боку голови.
Структурно-функціональна організація вестибулярної сенсорної системи, її рецепторний, провідниковий і кірковий відділи. Сприйняття положення голови в просторі та напрямку руху.
Вестибулярна сенсорна система
Велике значення для орієнтування людини у просторі має вестибулярна сенсорна система. Вона інформує ЦНС про положення голови, її переміщення, як активні, так і пасивні.
Вестибулярний орган складається з двох частин: статолітового (отоліто-вого) апарату, утвореного маточкою і мішечком, і трьох півколових каналів. Він є філогенетично древнішим, ніж інші сенсорні системи, і виник для сприйняття такого складного впливу, як дія сил земного тяжіння (рис. 24).
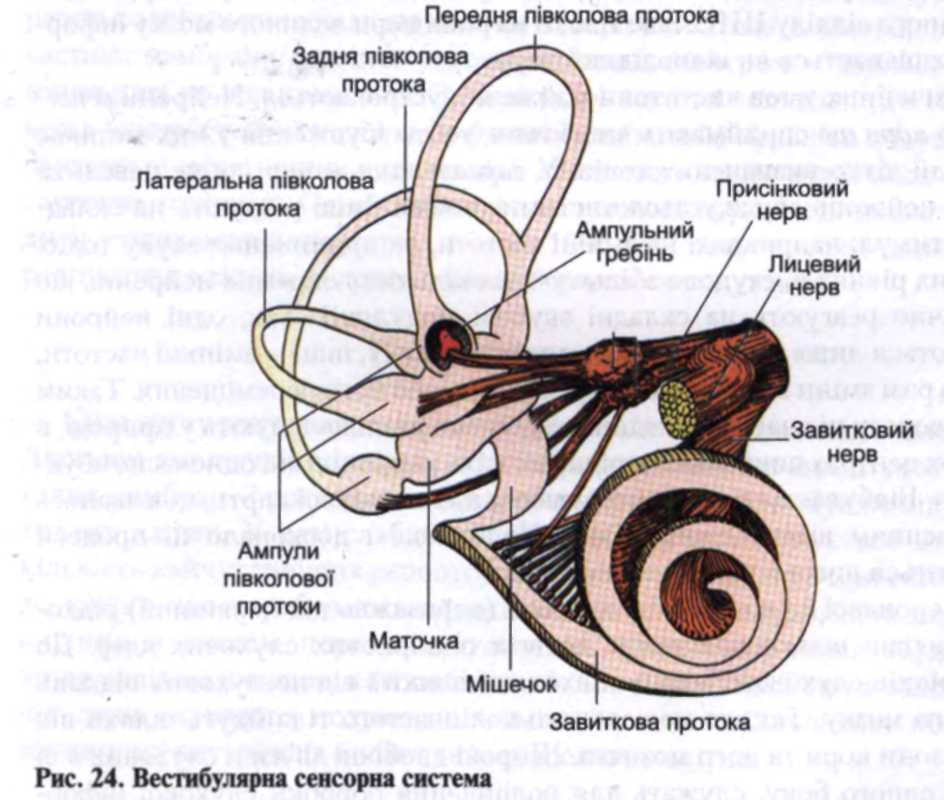
Рецептори вестибулярної сенсорної системи належать до вторинно-чутливих механорецепторів. У них перед чутливими нейронами є спеціальні рецептивні клітини, в яких під час дії відповідного подразника виникає збудливий рецепторний потенціал (ЗРП). На аферентний нейрон збудження передається за допомогою трансмітера - ацетилхоліну, що виділяється з цієї клітини.
Рецепторні клітини вестибулярного органу війчастого типу згруповані у п'ятьох місцях: у трьох ампулах півколових каналів і макулах (плямах) маточки та мішечка. Кожна клітина містить один найдовший рухомий волосок - кіноцилій і 50-60 коротших - стероциліїв. У разі згинання пучка коротких війок у бік кіноцилію у мембрані клітини підвищується проникність для №\ Це призводить до деполяризації - виникнення рецепторного потенціалу, внаслідок чого в синапс між клітиною та аферентним волокном виділяється трансмітер - ацетилхолін. Взаємодіючи з холіноре-цептором, він забезпечує виникнення генераторного потенціалу (ГП) в постсинаптичній мембрані аферентного волокна, який у разі сумації переходить у ПД. У зв'язку з тим що від рецепторних клітин вестибулярного аналізатора постійно відводяться ПД навіть за відсутності дії подразника, то під час нахилу війок у бік довгого волоска частота ПД збільшується. Водночас у разі нахилу війок у протилежний бік частота ПД знижується. Таким чином, в обох випадках ЦНС виявляється "обізнаною" про зміну впливу подразника.
Отолітовий апарат
Рецептори мішечка і маточки сприймають зміни положення голови у просторі, швидкість зміни прямолінійного руху і пов'язані з цим подразнення (вібрація, коливання тощо). У мішечку і маточці війки рецепторних клітин занурені у структуру отолітової мембрани, яка містить кришталики карбонату кальцію, у результаті чого питома щільність отолітової мембрани у два рази вища, ніж щільність ендолімфи. Сила інерції, що виникає під час лінійних прискорень на початку чи вкінці рухів "угору -вниз", "уперед - назад", діє на ендолімфу та отолітову мембрану по-різному. Важча, а отже, й інерційніша мембрана на початку руху відстає від руху ендолімфи, а під час гальмування пізніше зупиняється. У результаті саме в ці моменти і створюються умови для зсуву війок та виникнення збудження.
Отолітова мембрана маточки розташована у площині, близькій до горизонтальної, а мішечка - до вертикальної. У зв'язку з цим початок і закінчення горизонтальних рухів сприймаються переважно рецепторами маточки, а вертикальних - мішечка. Нахили голови також спричинюють "ковзання" мембрани, згинання війок і посилення деполяризації у відповідному відділі. Таким чином, за будь-якого положення голови кожна з отодітових мембран, займаючи певне положення щодо своїх рецепторних клітин, створює можливість для постійного його контролю. Рецептори мішечка найчутливіші до лінійних прискорень, які створюються під час вібрації.
Півколові канали
Природним стимулом для рецепторів півколових каналів є_кутаві прискорення. Тут рецептивні клітини кожного каналу згруповані в їхніх ампулах, утворюючи ампульні гребені. Кожен гребінь складається із волоскових та підтримувальних клітин, над якими розміщена желатиноподібна перегородка (купула), яка розділяє ампулу. Ці рецепторні утвори омиває ендолімфа, питома маса якої мало відрізняється від желатиноподібної маси, тому війкові клітини цих відділів майже не реагують на лінійні прискорення.
Перетинчастий лабіринт кожного каналу завдяки наявності загальної частини утворює замкнуте, хоча й не зовсім ідеальне коло. Тіло рецепторної клітини і її війки під час обертального руху голови перебувають у різних умовах. Рідина (ендолімфа) через інерцію на початку руху залишається ще якийсь час нерухомою, а півколові канали разом з рецепторами вже рухаються. Тому виникає різниця тисків по обидва боки купули. У результаті купула відхиляється у протилежний руху бік, війки згинаються, що призводить до виникнення рецепторного потенціалу. Збудження у рецепторах виникає як на початку, так і вкінці обертальних рухів голови, а також під час зміни швидкості обертання. За рівномірної швидкості обертання тіло клітини і війки рухаються разом, і в цей час подразнення рецептора відсутнє.
У зв'язку з тим що півколові канали лежать у трьох площинах (хоча передня частина горизонтального каналу піднята приблизно на 30°), рецептори кожного з них сприймають зміни руху навколо відповідної осі обертання. У тому разі, якщо площина обертання розташована між півколовими каналами (не збігається з одним із них), то збуджуються рецептори різних ампул з різною інтенсивністю. Тому ми можемо сприймати зміну швидкості обертання у всіх площинах.
Центральні відділи вестибулярної системи
Збудження від рецепторних клітин синаптично передається до дендритів нейронів присінкового вузла, що лежить на дні внутрішнього слухового каналу. Аксони цих нейронів утворюють присінкову частину VIII пари черепних нервів і підходять до вестибулярних ядер моста мозку. Аксони цих ядер йдуть до ядер мозочка і спинного мозку. Частина волокон від вестибулярних ядер іде додаламуса, а звідти імпульси прямують до пдстч^+тральщд.звивини кори великого мозку. Крім того, нейрони вестибулярних ядер контактують з багатьма іншими відділами ЦНС: із ядрами III, IV, VI, X пар черепних нервів, ретикулярною формацією, гіпоталамусом. Це зумовлює залучення багатьох функціональних систем організму до відповіді на подразнення вестибулярних рецепторів. У разі інтенсивного подразнення рецепторів вестибулярної системи виникають моторні рефлекси, очний ністагм (спочатку відбувається повільний поворот очного яблука в бік, протилежний обертанню, а потім швидкий рух у бік обертання, коли око ніби доганяє обертання голови), вегетативні розлади (зміна ЧСС, звуження судин шкіри, потовиділення, нудота та інші прояви, характерні для "морської хвороби").
У результаті поєднання активності нейронів вестибулярного нерва (це переважно функція отолітового апарату) і рухових ядер стовбура та мозочка зароджується багато моторних рефлексів, спрямованих на підтримання пози (положення).
Імпульси від вестибулярної системи, які проходять через таламус, надходять до скроневої ділянки кори великого мозку, де забезпечується аналіз отриманої інформації, а отже, і свідоме орієнтування у просторі.
ЗАКЛЮЧЕННЯ.
Підводячи підсумки лекції, можна зробити такі висновки :
1 – для гарного розуміння процесів нервової регуляції діяльності організму треба добре знати фізіологію ЦНС,
2 –всі відділи ЦНС тісно пов`язані між собою анатомічно і фізіологічно,
3 – знання особливостей фізіологічних процесів в нормі дозволить більш глибоко зрозуміти ті зміни, які відбуваються в організмі при хворобах, що для медичного працівника має велике значення.
А тепер для того , щоб з`ясувати , наскільки добре ви засвоїли матеріал лекції проведемо невелике тестування .
А. Тести для самоконтролю:
Із запропонованих відповідей виберіть правильну .