

Типи рухливості окремих елементів мембрани:
а – латеральна ( бічна) рухливість ліпідів, б – обертальні рухи, в – латеральна рухливість білків, г – « фліп – флоп» ліпідів, д – « фліп – флоп» білків.
Таким чином, для подвійного шару фосфоліпідів характерною є постійна зміна стану з високою рухомістю його компонентів. Рухомість мембрани полегшує процеси транспорту речовин через неї.
Фосфоліпідні фракції, що підтримують клітинні мембрани у стані рідкої фази, в різних умовах життєдіяльності можуть змінюватися. Так, наприклад, з появою великої кількості гормонів, що характерно для стресу та інших подібних станів, може зростати швидкість утворення гідроперекисів ліпідів, які підсилюють сегментарну рухомість фосфоліпідів. Це істотно змінює властивості мембран, зокрема їх проникність. А за умови старіння відбувається нагромадження вільних радикалів і знижується синтез фосфоліпідів, що підтримують мембрани у стані рідкої фази.
Бімолекулярний шар ліпідів є матриксом, каркасом мембрани, в який занурені білкові молекули, що утворюють канали для води і йонів, формують йонні насоси тощо (мал. 109, а).
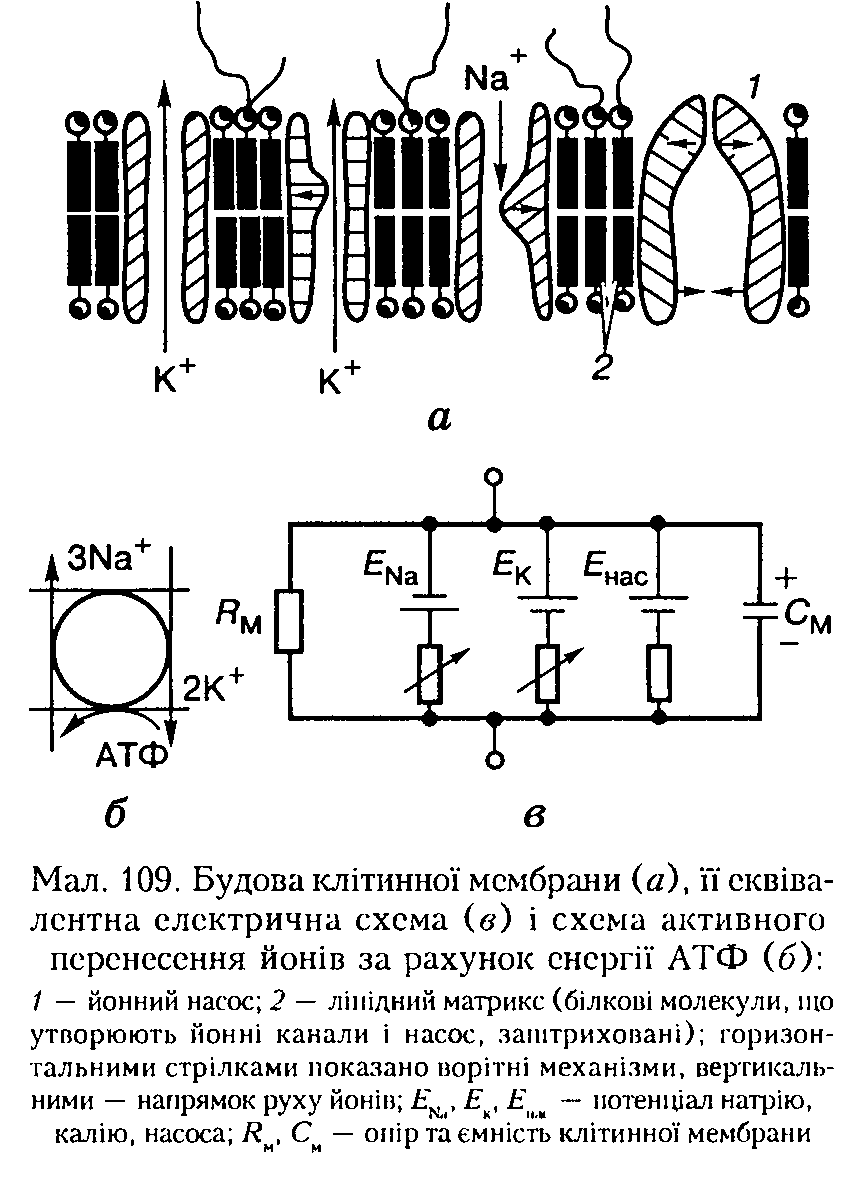
Окремі фрагменти молекул фосфоліпідів чинять тепловий рух, що відіграє важливу роль:
1) у взаємодії ліпідів з білками;
2) у процесах транспорту речовин через мембрану;
3) у проникності (рис. 2).
Підвищенню рухливості мембрани сприяє наявність у молекулі ліпідів коротколанцюж-
кових жирних кислот і розгалужених ланцюгів. Чим вищий ступінь їхньої ненасиченості, тим сильніше виражений ступінь розрідженості мембрани.
Ферментативна активність і пасивний транспорт тісно пов’язані з рухливістю мембранних ліпідів.
Білки мембран. Ззовні і зсередини до ліпідів прилягають два білкові шари Частка білків становить близько 55% речовини мембран.
Але, на відміну від ліпідів, білки не скрізь утворюють суцільні шари.
Час життя білків становить від двох до п'яти днів, тому в клітині на полісомах йде постійний синтез білків мембран.
Більшість функцій мембран зумовлені їх білковими компонентами.
Білки виконують функції:
іонних каналів,
помп,
ензимів,
рецепторів.
Активність і функції білків залежать як від них самих та їх щільності на мембрані, так і від ліпідів мембран.
Білки мембрани бувають двох видів:
інтегральні,
периферійні.
Інтегральні білки пронизують мембрану наскрізь. Вони можуть входити до складу іонних каналів, виконувати роль ензимів та переносників через мембрану великих молекул. Більшість з них за своїм хімічним складом є глікопротеїнами.
Їхня вуглеводна частина виступає з клітинної мембрани назовні і може бути носієм антигенів або рецептором, що зв'язується з молекулами гормонів.
Глікопротеїни утворюють також адгезивні молекули, які сприяють прикріп-
ленню клітин одна до одної.
Інтеґральні мембранні білки при фізіологічній температурі також здійснюють різні рухи ( див мал.).
Периферійні білки вбудовані на різну глибину між ліпідами. Вони локалізовані на поверхні мембрани і зв'язані з нею переважно слабкими електростатичними силами.
Периферійні білки переважно представлені ензимами (ацетилхолінестераза, фосфатази тощо).
Так як і полярні ліпіди, периферичні білки мають одну полярну область і одну неполярну (домен).
У полярній області білкової глобули, що знаходиться в контакті з водою, зібрані іонізовані залишки амінокислот і всі ковалентно зв’язані вуглеводні залишки.
У неполярній області відсутні іонізовані і вуглеводні залишки. Ця область білкової глобули занурена в гідрофобну внутрішню частину мембрани
За функціональними особливостями мембранні білки всіх клітин ділять на 5 класів ( див. рис 2.1):
- білки-насоси,
- білки-канали,
- білки-рецептори,
- ферменти і
- структурні білки.
Білки-насоси витрачають метаболічну енергію АТФ для переміщення іонів і молекул проти концентраційних і електрохімічних градієнтів і підтримують необхідні концентрації цих молекул у клітині.
Білки-канали являють собою шляхи вибіркового перенесення іонів і заряджених молекул. Механізм перенесення пов'язаний з конформацією білка-каналу, в результаті якої він відкривається або закривається. Багато білків-каналів зв'язані з білками - рецепторами .
Білки-ферменти: володіють високою каталітичною активністю, полегшують або уповільнюють протікання хімічних реакцій як у самій мембрані, так і на її поверхні.
Структурні білки: забезпечують поєднання клітин у тканини і органи. Вони також відіграють важливу роль у підтриманні разом з білками цитоскелета форми клітин.
Скорочувальні білки, що мають АТФазну активність, тобто здатність розщеплювати АТФ із утворенням АДФ і фосфату, беруть участь у процесі екзоцитозу.
Білки-рецептори ( є периферичними білками) представлені білковими молекулами, які "впізнають" ту чи іншу біологічно активну речовину – ліганд ( гормон, БАР, тощо), за допомогою складних біохімічних перетворень контактують з ним, кодують і передають інформацію про характер цих взаємодій у клітину. Від характеру взаємодії рецептора з лігандом значною мірою залежить кінцевий фізіологічний ефект дії останнього.
Рецептор може бути самостійною структурою, вбудованою у мембрану у вигляді інтегрального білка, або частиною інших функціональних білків і регулювати їх активність. Для одного і того ж хімічного агента на мембрані може бути кілька рецепторів й ефект взаємодії субстрату з ним не тільки не подібний, а й у деяких випадках навіть діаметрально протилежний. Так, у разі взаємодії гормону мозкового шару надниркових залоз адреналіну з А-адренорецептором спостерігається звуження кровоносної судини, а з В-рецептором - розширення.
Рецептори мембрани здатні сприймати хімічні або електричні сигнали. Результатом цього є хімічна модифікація мембранних білків й утворення "посередників" - месенджерів - речовин, що забезпечують зміну швидкості та напрямку біохімічних процесів у клітині , а значить, зміну її функції.
Такими посередниками( месенджерами) можуть бути:
- іони кальцію.