

Характеристика натрієвих, калієвих та кальцієвих каналів.
НАТРІЄВІ КАНАЛИ.
Натрієвим каналам властива висока, але не абсолютна вибірковість до Na+, оскільки більшою або меншою мірою вони є проникними для йонів літію, талію і деяких органічних катіонів( гідразин, амоній)
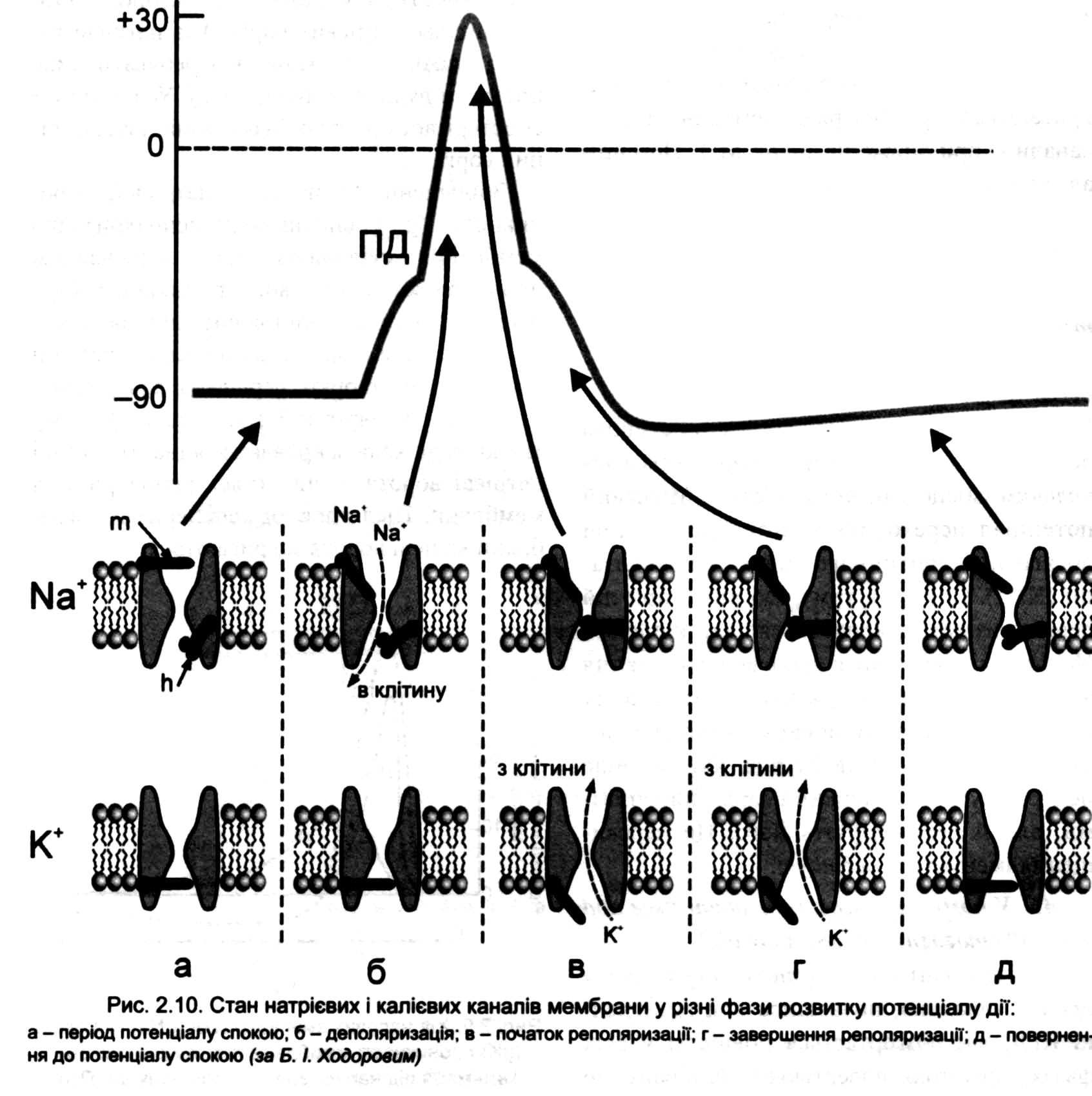
Через потенціалозалежні Nа+канали в клітину за концентраційним і електричним інградієнтами входять іони Nа+, кожний з яких привносить значний позитивний заряд. Розвивається подальша швидка часткова зміна мембранного потенціалу.
Ця деполяризація відкриває додаткову кількість тих самих каналів з повторенням усього процесу: вхід Nа , деполяризація клітинної мембрани - відкриття швидких потенціалзалежних натрієвих каналів і т. д. Це фаза швидкої деполяризації мембрани.
Весь процес повторюватиметься доти, поки не відкриються всі такі канали. За суттю процес нагадує вибух атомної бомби внаслідок ланцюгової реакції, через що й має назву - авторегенераторного, лавиноподібного.
У момент відкриття всіх швидких потенціалзалежних натрієвих каналів вхідний натрієвий струм стане максимальним і відбудеться перезарядка мембрани (реверсія поляризації, овершут або "переліт") - внутрішня поверхня мембрани клітини зарядиться позитивно, зовнішня - негативно.
Натрієві канали у відкритому стані можуть бути дуже короткий час, і їх активація змінюється інактивацією (закриваються інактиваційні h-ворота - схожу будову мають і кальцієві канали.
Вхідний струм зменшується, розвивається реполяризація мембрани (відновлюється її попередній заряд).
Процес прискорюється тим, що до початку реполяризації відкриваються і калієві канали (їх також відкриває зміна потенціалу мембрани при подразненні). У результаті посилюється вихід К+.
На мал. показано зміни натрієвої і калієвої провідності мембрани нервової клітини під час фіксованої деполяризації мембрани. Рівень провідності цих йонів відбиває кількість одночасно відкритих натрієвих і калієвих каналів. Як можна легко побачити, що рівень провідності натрію за частки мілісекупди досягає максимуму, а потім повільно знижується до початкового рівня. Це пояснюється наявністю у натрієвих каналах двох типів воріт — швидких активаційних і повільних інактиваційних.
Калієві канали
Калієві канали за будовою подібні до натрієвих, але відрізняються своєю вибірковістю. В усіх збудливих клітинах калієві канали відповідають за пізній йонний струм. Діаметр калієвого каналу у найвужчій його частині становить 0,3 нм. Отже, калієві канали проникні для катіонів розміром 0,26-0,З нм.
Активація цих каналів порівняно з натрієвими відбувається досить повільно. Крім того, протягом перших 10 мс деполяризації немає ніякої калієвої інактивації, вона виникає тільки під час дуже тривалої (кілька секунд) деполяризації мембрани.
У мембрані багатьох нервових і м'язових клітин існують калієві канали, які інактивуються порівняно швидко.
Калієві канали мають тільки одні активаційні ворота. Пізнє відкриття цих каналів обумовлено більш повільною кінетикою їх активації.
Щільність розміщення калієвих каналів на мембрані дещо менша, ніж натрієвих. Специфічним блокатором калієвих каналів є тетпраетиламоній (ТЕА), йони гідрогену, амінопіридини. ТЕА діє з будь-якого боку клітинної мембрани.
Відкриття натрієвих каналів можна порівняти з відкриттям маленької хвіртки на пружині, калієвих - важких воріт на громіздких завісах.
Відповідно і натрієвий струм буде швидко розвиватися та згасати, а калієвий - більш повільно досягати максимуму й поступово зменшуватися. Просвіт каналу - це селективний фільтр для певних іонів.
КАЛЬЦІЄВІ КАНАЛИ.
Виявилося, що канали Са2+ також беруть активну участь у генерації ПД.
В деяких клітинах вхідний струм створюється переважно Са2+ (гладком'язові клітини).
Кальцієві канали мають дещо уповільнену (порівняно з натрієвими каналами) кінетику процесу активації (мілісекунди) і ще повільнішу кінетику процесу інактивації (десятки і сотні мілісекунд).
Селективність (вибірковість) кальцієвих каналів зумовлена підвищеною спорідненістю до двовалентних катіонів: йони кальцію зв'язуються з особливими хімічними групами і тільки після цього проходять до порожнини каналу.
До деяких двовалентних катіонів, наприклад Мп2+, спорідненість цих хімічних груп така велика, що, зв'язуючись з ними, ці катіони блокують рух Са2+ через канал. Специфічними блокаторами кальцієвих каналів є харибдотоксин, вилучений з отрути скорпіона, а також такі лікарські речовини, як верапаміл та ніфедипін. Останні широко застосовуються при деяких серцево-судинних хворобах.
Характерною особливістю кальцієвих каналів є залежність їх від клітинного метаболізму, зокрема від циклічних нуклеотидів (цАМФ, цГМФ), які регулюють процеси фосфорилування і дефосфорилування білків кальцієвих каналів. Йони кальцію ефективно впливають на синтез білків і забезпечують транспорт утворених молекул до аксонів і дендритів. Відкриття цього явища безпосередньо свідчило про те, що процеси на мембрані прямо пов'язані з процесами всередині клітини.
Дослідження останніх років показали, що Са2+ є важливим регулятором активності збудливих систем. Переважна кількістьСа , що потрапляє в клітини під час збудження зв’язується буферними системами, а вільна частина бере активну участь у у таких процесах, як регуляція функції К –каналів обміну кальцію та натрію.
Са2+-помпа.
Існує два типи кальцієвої помпи:
- один забезпечує викид іонів із клітини в міжклітинне середовище,
- інший переносить іони із цитоплазми усередину саркоплазматичного ретикулуму м'язових волокон, де створюється депо кальцію.
Енергетична ємність цієї помпи набагато більша, ніж ємністьNa+-К+-АТФ-ази: для викачування одного іона Са2+ витрачається дві молекули АТФ, у той час як одна молекула АТФ витрачається для транспорту трьох іонів Na+двох іонів К+. Регулює роботу цієї помпи сам кальцій.
Найменша зміна вутрішньоклітинної концентрації Са2+ запускає механізм його відкачування.
Виявлено ще одну особливість кальцієвих каналів: вони здатні дуже специфічно реагувати на деякі фізіологічно активні речовини, що утворюються іншими клітинами і виділяються у внутрішнє середовище організму (катехоламіни, пептиди тощо). Під впливом цих речовин канали активуються чи, навпаки, інактивуються. Внаслідок цього змінюється активність нервової клітини.