

IV. За швидкістю відкривання чи закривання ворота:
1) швидкі
2) повільними.
Так, активаційні ворота натрієвих каналів є швидкими, тобто відкриваються відразу під час дії подразника. Інактиваційні ворота натрієвих каналів і ворота калієвих каналів - повільні, тобто відкриваються і закриваються повільніше.
V. За розміром селективної ділянки:
1) селективних,
2) неселективних.
Селективні пропускають лише якийсь певний йон: №+, К+ або Са2+, а неселективні — кілька різних йонів.
Vі. В залежності від виду контролю за каналом:
1) потенціалозалежні,
потенціалонезалежні (хемочутливі) і
канали витоку.
Потенціалозалежні канали контролюються рівнем МПС. Потенціалонезалежні (хемочутливі) канали контролюються тільки хеморецепторами. Канали витоку також потенціалонезалежні, але не контролюються хеморецепторами, мають значення в потенціалі спокою.
Розглянемо більш детально останню класифікацію . Потенціалозалежні канали.
Потенціалозалежними є натрієві, калієві і кальцієві канали. Вони забезпечують первинно-активний транспорт іонів
Провідність цих каналів контролюється рівнем МПС, тобто ці канали відкриваються (активуються) і закриваються (інактивуються) у відповідь на зміни МПС. Потенціалозалежні канали функціонують в електрозбудливих мембранах і беруть участь у генерації ПД. Електрична збудливість повязана з існуванням у таких каналах особливого молекулярного пристрою —воріт, відкривання і закривання яких визначається електричним полем мембрани.
Ворота можуть перебувати тільки у двох положеннях:
повністю відчинених
чи повністю зачиненими,
тому провідність поодинокого йонного каналу є сталою.
Відкриття, закриття та інактивація воріт залежать від мембранного потенціалу.
Вузька частина каналу, що є селективним фільтром, дуже коротка і має одиничний негативний заряд. Цей заряд притягує катіони, відштовхує аніони, забезпечуючи вибіркову проникність для Na+ і подібних до нього катіонів.
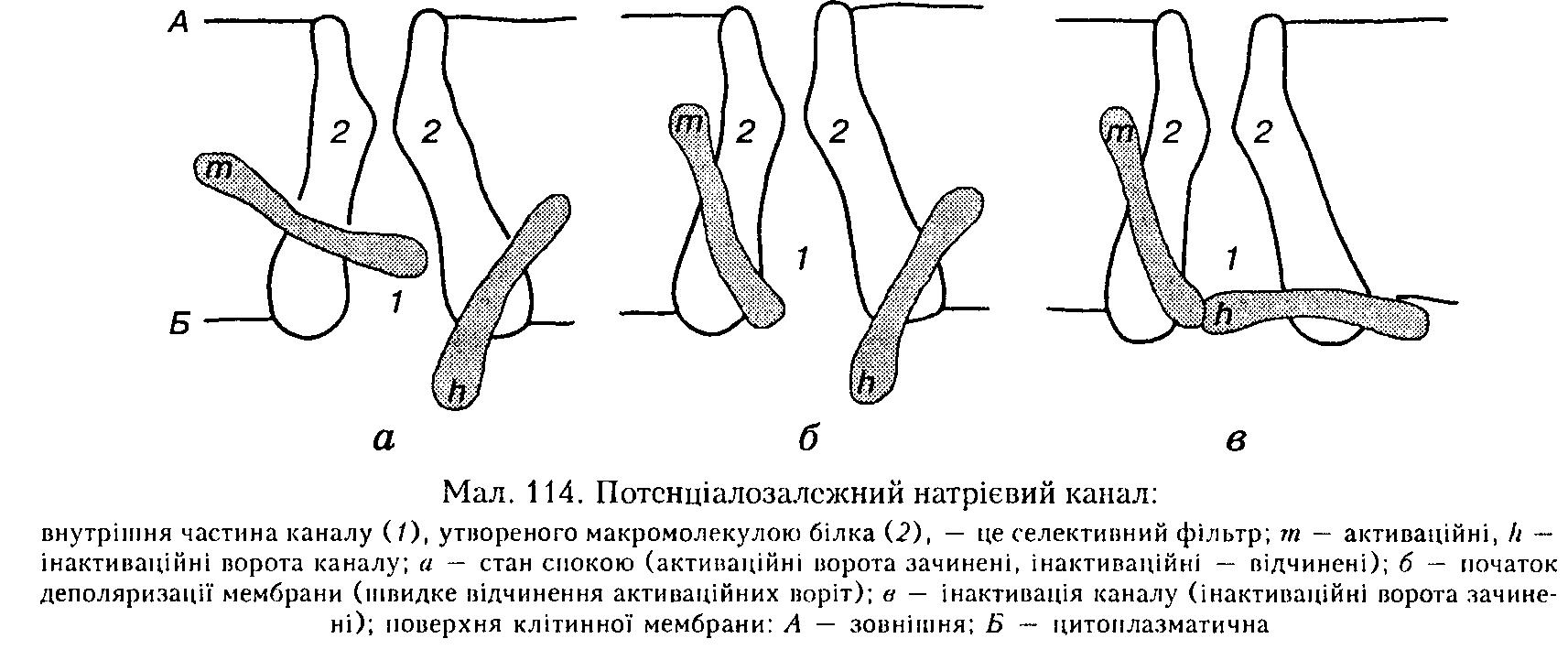
Активаційні та інактиваційні ворота розміщені біля внутрішнього кінця натрієвого каналу, причому інактиваційні ворота спрямовані в бік цитоплазми (мал. 114).
У стані спокою активаційні ворота закриті, а інактиваційні відкриті. Під час деполяризації мембрани спочатку відкриті обидва типи воріт, тобто канал пропускаєNa+. Потім інактиваційні ворота повільно закриваються — канал інактивується. Лише після закінчення деполяризації поступово відкриваються інактиваційні ворота, а активаційні закриваються і канал повертається до початкового стану.
Деякі речовини специфічно змінюють процеси активації та інактивації натрієвих каналів. Так, батрахотоксин спричинює стійку деполяризацію клітинної мембрани, підвищуючи проникненість натрію Він повністю усуває процес натрієвої інактивації так, що натрієві канали стають постійно відкритими. Тетродотоксин дуже швидко і різко пригнічує ранній йонний струм крізь мембрану, але тільки тоді, коли вій діє іззовні, тобто на чутливі до тетродотоксипу рецепторні структури натрієвих каналів, розміщені на зовнішній поверхні мембрани.
Через відкриті канали йони рухаються за концентраційним і електрохімічним градієнтами. Ці потоки йонів зумовлюють зміну МПС, що в свою чергу, змінює середню кількість відкритих каналів і відповідно рівень йонних струмів.
Для дослідження функції йонних каналів застосовують різні методи. Один із них -це метод фіксації напруги на мембрані, за допомогою якого на мембрані штучно підтримується будь-який потрібний потенціал.