

Гальмівні синапси, їх нейромедіатори.
Гальмування — активний нервовий процес, який призводить до зменшення чи припинення збудження в локальній ділянці нервової тканини.
Вперше гальмування в ЦНС описав І. М. Сєченов (1862). Завдаючи хімічного або слабкого електричного подразнення па середній мозок жаби, він спостерігав гальмування її згинального рефлексу, що виявлялося у зростанні часу цього рефлексу — часу від початку подразнення рецептора до виникнення відповідної реакції.
Завдяки цьому відкриттю було доведено існування у центральній нервовій системі структур і механізмів, що гальмують рефлекторну діяльність нервових центрів, розміщених нижче, і показано виняткову роль їх у координації будь-яких рефлексів.
Відомо, що гальмування — це активний нервовий процес, який викликається збудженням і виявляється пригніченням іншого збудження.
На відміну від процесу збудження, гальмування може розвиватися тільки у вигляді локального процесу і тому завжди пов'язане з існуванням спеціальних гальмівних синапсів.
Гальмівними нейронами є:
- у спинному мозку є вставні нейрони — клітини Реншо,
- в мозочку — грушоподібні нейрони (Пуркіньє).
Розрізняють кілька видів гальмування у ЦНС:
За механізмами виникнення воно поділяється на:
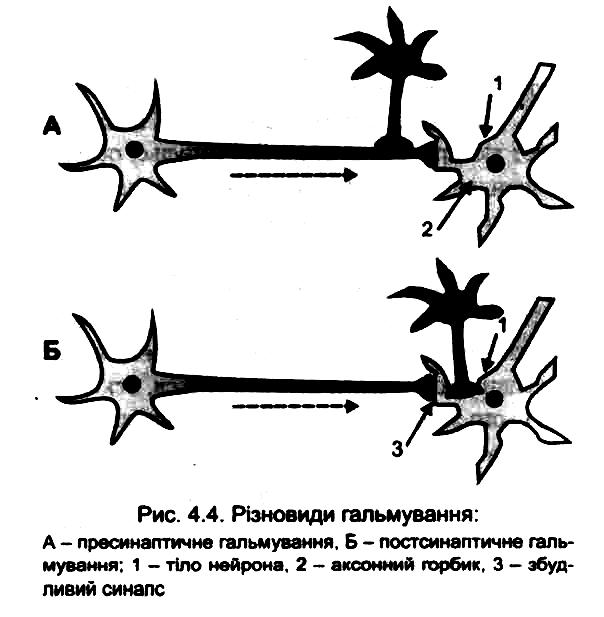
постсинаптичне (розвивається на тілі нейрона) і
пресинаптичне (розвивається на аксоні нейрона перед його синаптичним закінченням);
За механізмом розвитку поділяється на:
гіперполяризаційне і
деполяризаційне;
За напрямком поширення і будовою нейронних ланцюгів – на:
пряме,
поєднане,
латеральне,
реципрокне та
зворотне.
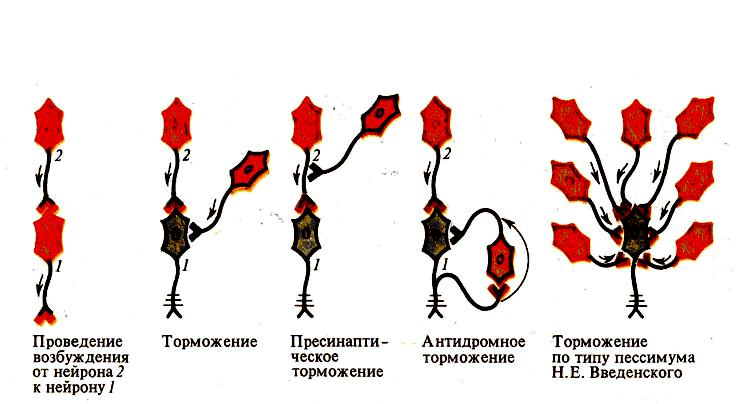
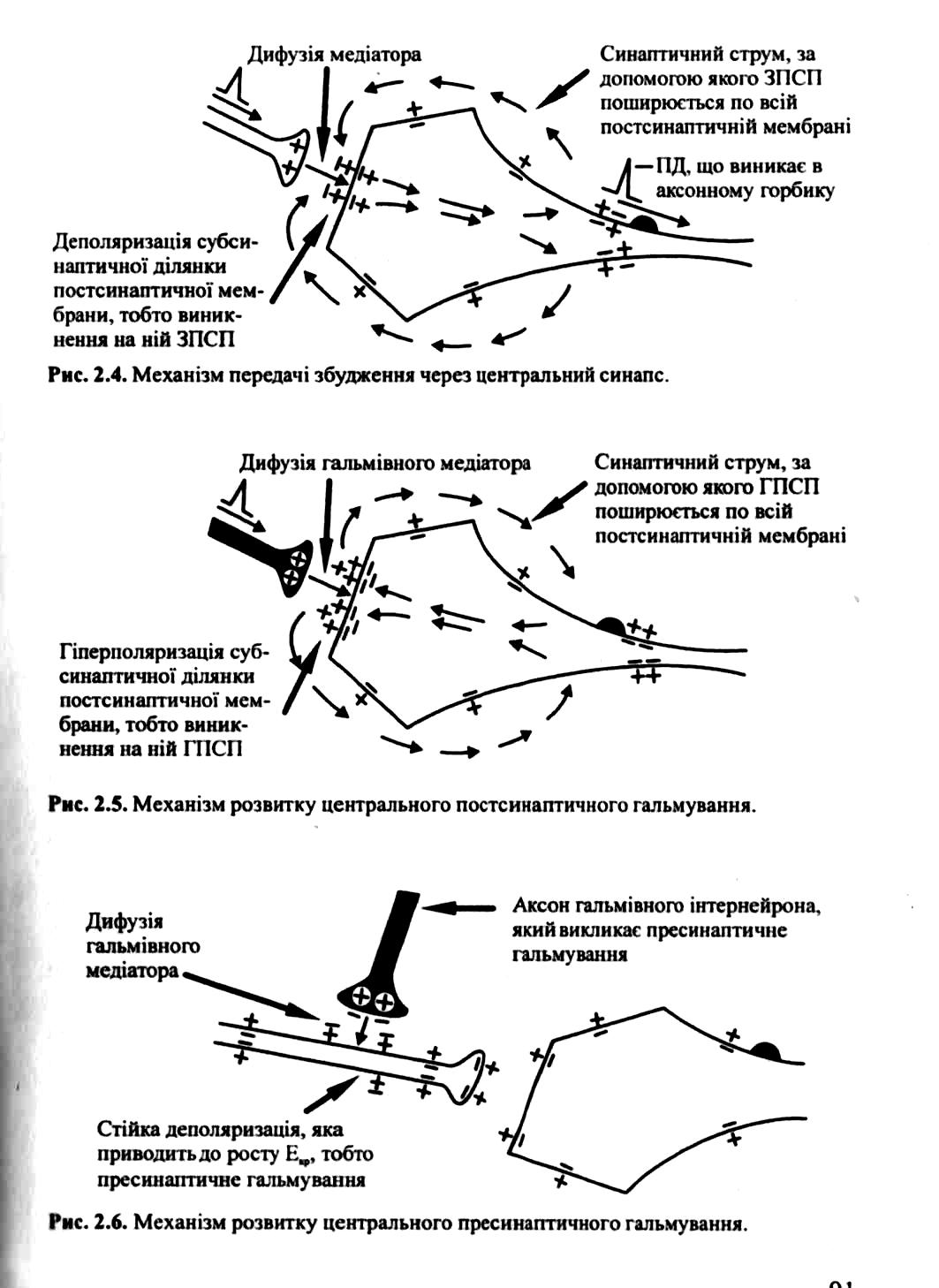
1. Постсинаптичне гальмування є найпоширенішим у ЦНС. Воно полягає в тому, що медіатор, який виділяється пресипаптичним закінченням гальмівних нейронів, змінює властивості постсинаптичної мембрани таким чином, що вона не може генерувати збудження.
В основі цього виду гальмування лежить гальмівний постсинаптичний потенціал (ГПСП), який є локальною гіперполяризацією мембрани, що виникає внаслідок підвищення її проникності до Са або К+. Постсинаптичне гальмування може бути прямим і зворотним.
А) Пряме постсинаптичне гальмування виникає у постсинаптичному нейроні внаслідок гіперполяризації його мембрани під час розвитку ГПСП під впливом ПД будь-якого гальмівного нейрона.
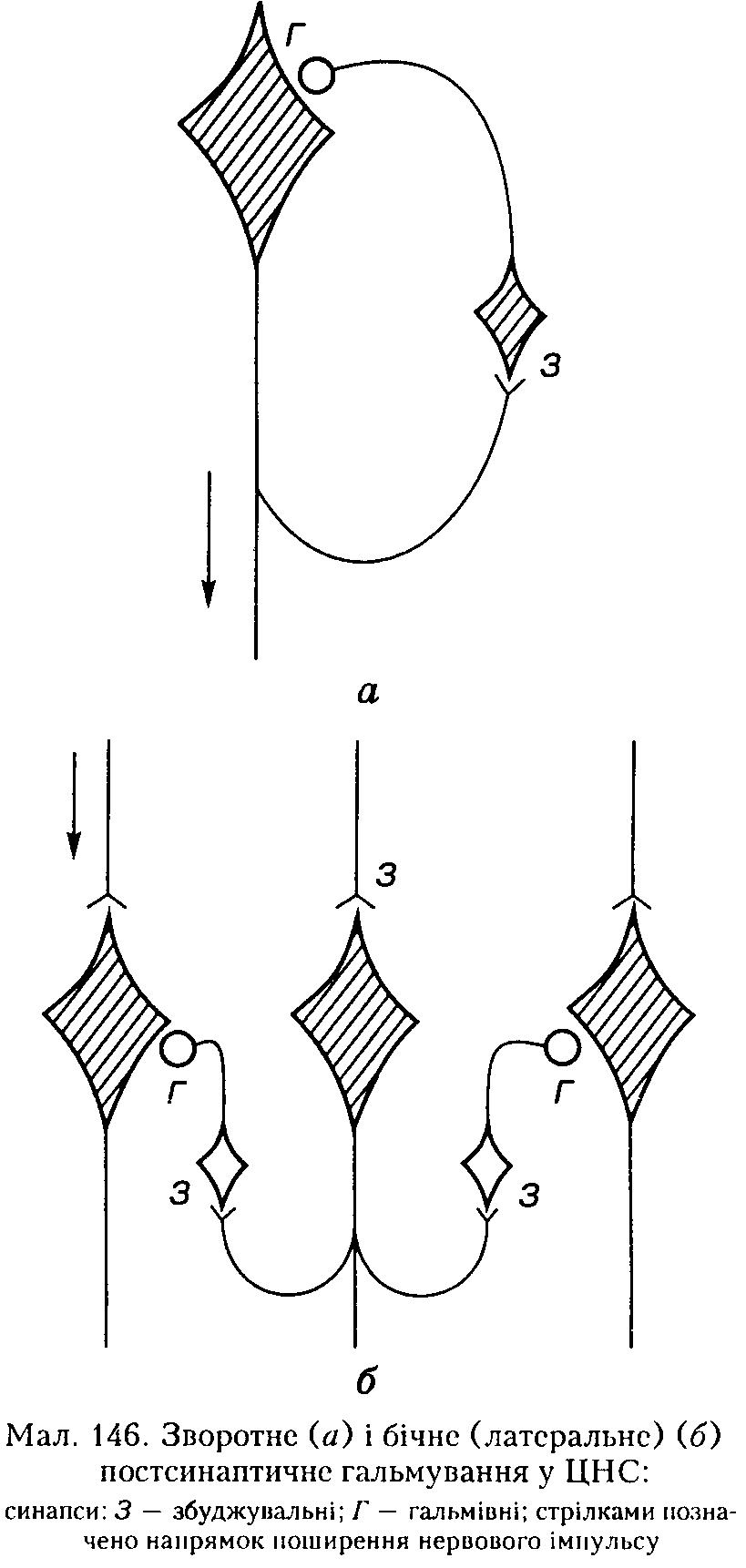
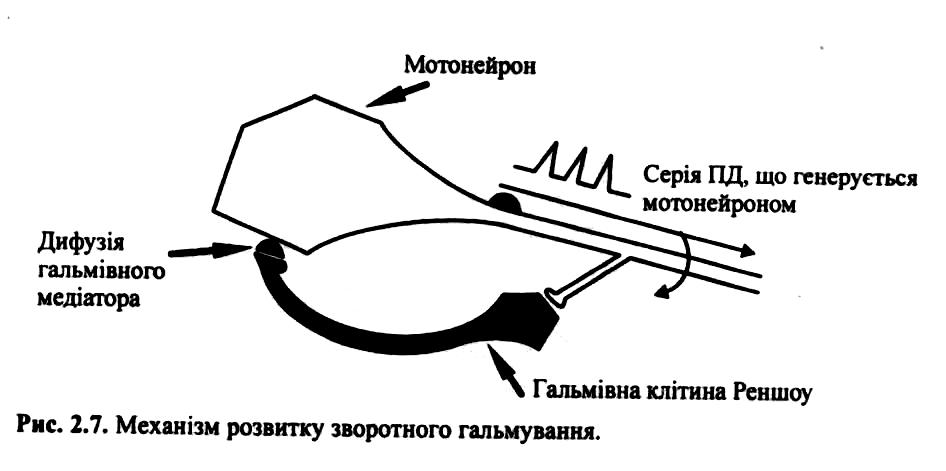
Б) Зворотне постсинаптичне гальмування здійснюється у нейронному ланцюзі, який складається з рухового нейрона та вставного гальмівного нейрона (клітини Реншо). Імпульси від збудженого рухового нейрона не тільки прямують через аксон, а й поширюються через його колатералі, що відгалужуються від нього, і активують вставний нейрон — клітину Реншо.
Цей гальмівний нейрон спричинює пригнічення розрядів рухового нейрона (мал. 146, а). Отже, з цих двох нейронів утворюється контур з негативним зворотним зв'язком, який дає змогу стабілізувати частоту розрядів рухового нейрона, обмежуючи надмірну імпульсацію до ефекторпого органа.
Іноді вставні нейрони формують гальмівні синапси не тільки на рухових нейронах, що активують своїм збудженням саме ці клітини, а й на суміжних рухових нейронах, які виконують подібні функції. Таке гальмування суміжних нейронів називають бічним (латеральним) (див. мал. 146, б).
Ефективність гальмівних синапсів багато в чому залежить від їх локалізації на поверхні нейрона:
гальмівний ефект тим більший, чим ближче розміщений гальмівний синапс до аксонного горбка, бо саме там відбувається генерація ПД.
У нейронах кори великого мозку і таламуса виявлено два види постсинаптичного гальмування:
1) соматичне, яке виникає під час збудження гальмівних синапсів, розміщених на тілі і найближчих дендритах нейрона внаслідок генерації високоамплітудних ГПСП, що блокують реакції нейрона на будь-яку аферентну імпульсацію;
2) дендритне, яке з'являється під час збудження гальмівних аксодендритних і дендро-дендритних синапсів, розміщених на дистальних ділянках дендритів. ГПСП, що виникають при цьому, завдяки своїй невеликій амплітуді і значній відстані між місцем виникнення і аксоним горбком не здатні викликати значні зміни МП нейрона. Отже, дендритне гальмування має локальний характер і забезпечує вибіркове пригнічення аферентного входу до нейрона через певні частини дендритного дерева, відіграючи важливу роль в інтегративній діяльності мозку (П. М. Сєрков).
 Постсинаптичне
гальмування
- це
основний вид гальмування у ЦНС. Типовим
гальмівним синапсом є аксо-соматичний
синапс.
Постсинаптичне
гальмування
- це
основний вид гальмування у ЦНС. Типовим
гальмівним синапсом є аксо-соматичний
синапс.
Основними трансмітерами, що спричиняють цей вид гальмування, є ГАМК та гліцин.
Постсинаптичне гальмування значно поширене в ЦНС. Воно реалізується у нервових центрах, у мотонейронах спинного мозку, в симпатичних гангліях.
2. Пресинаптичне гальмування також здійснюється за допомогою спеціальних вставних нейронів, проте механізм його інший і досить складний. Структурною основою цього гальмування є аксоаксонні синапси, тобто аксон проміжного гальмівного нейрона утворює додатковий синапс на аксоні збуджувального нейрона. Гальмування розвивається завдяки зменшенню або блокаді виділення трансмітера в синаптичну щілину збуджувального синапса, тобто гальмування передачі нервових імпульсів відбувається завдяки зміні властивостей його пресинаптичної мембрани. Гальмування не поширюється на постсинаптичну мембрану такого синапса. (мал. 147).
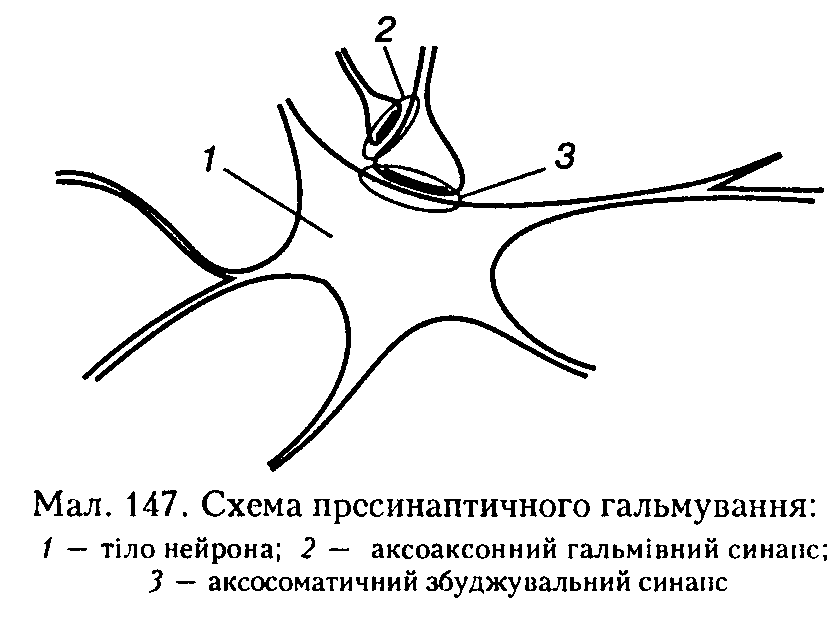
Медіатор, що виділяється в цих синапсах (припускається, що це ГАМК), викликає деполяризацію нервових закінчень за рахунок збільшення проникності мембрани для Сl. Іншою можливою причиною такої деполяризації може бути підвищення зовнішньої щодо нейрона концентрації К+ при тривалій активації аферентних входів. Гальмування збуджуваного нейрона виникає тому, що ця пресинаптична деполяризація спричинює зменшення амплітуди ПД у пресинантичному закінченні, що, в свою чергу, призводить до зменшеного виділення квантів збуджувального медіатору (нейротрапсмітера), тому амплітуда ЗПСП знижується, і активація збуджуваного нейрона не відбувається.
Конкретний механізм пресинаптичного гальмування з'ясований недостатньо. Вважають, що трансмітер, який вивільняється в аксоні гальмівного нейрона, спричиняює стійку, тривалу деполяризацію закінчень збуджувальних нейронів. Протягом цього часу розвивається інактивація натрієвих каналів, унаслідок чого знижується збудливість мембрани аксона збуджувального нейрона в ділянці гальмівного синапса. Тому потенціали дії або не доходять до збуджувального синапса, або їх амплітуда знижується і кількість вивільненого збуджувального трансмітера стає недостатньою для генерації ПД у нейроні, де відбувається процес гальмування.
Можливий також інший механізм. Гальмівний трансмітер може зменшувати проникність пресинаптичної мембрани збуджувального синапса щодо іонів кальцію. Це призводить до зменшення викиду трансмітера в синаптичну щілину цього синапса, амплітуда ЗПСП у нейроні зменшується і він гальмується.
Основним трансмітером для цього виду гальмування є ГАМК, а пригнічують його такі антагоністи, як бікукулін, пікротоксин (судомні отрути). У разі їх надходження в організм розвиваються судоми, оскільки велика кількість таких синапсів локалізована на мотонейронах спинного мозку.
Пресинаптичне гальмування значно триваліше (сотні мілісекунд) за постсинаптичне.
Воно особливо ефективне для обробки інформації, яка надходить до нейрона різними пресинантичними шляхами, оскільки в цьому разі збудження, що надходить через один із входів, може бути вибірково зменшене або навіть повністю пригнічене, якщо немає інших виливів на інші синаптичні входи. Припускають, що пресинаптичне гальмування пригнічує сигнали, біологічне значення яких у даний момент є незначним, і пропускає інформацію, що є найважливішою для організму.
Вважають також, що пресинаптичне гальмування відсікає маловажливі аферентні сигнали, постсинаптичне - узгоджує між собою діяльність антагоністичних нервових центрів.
Відмінності цих двох видів гальмування полягають у тому, що
пресинаптичне гальмування вибірково вимикає окремі входи до нервової клітини, тобто відсікає маловажливі аферентні сигнали,
постсинаптичне гальмування знижує збудливість нейрона в цілому; узгоджує між собою діяльність антагоністичних нервових центрів.
Таким чином, локальні процеси гальмування, незважаючи на те, що не здатні до поширення, блокують проходження збудження, обмежують його поширення, запобігаючи цим перезбудженню ЦНС.
Песимальне (лат. — найгірший) гальмування. Про нього вже йшлося під час розгляду передачі збудження у нервово-м'язовому синапсі (див. с. 279). М. Є. Введенський вважав, що песимальне гальмування виникає внаслідок зниження функціональної лабільності збудливої структури в разі надмірних за силою або частих подразнень і виявляється пригніченням рефлекторної діяльності. Песимальне гальмування відіграє важливу роль у механізмах координації рефлекторної діяльності в ЦНС. Воно не пов'язане з активацією спеціалізованих гальмівних утворів, а виникає внаслідок поступового зниження збудливості, і ті самі імпульси, що раніше викликали рефлекторний акт, тепер його гальмують.
3) Реципрокне гальмування, як один із видів складної координації рефлекторної діяльності
Реципрокне (лат.— взаємний), спряжене (поєднане) гальмування є важливим механізмом координаційної діяльності. Якщо подразнювати кінцівку спінального собаки електричним струмом, виникає звичайний згинальний рефлекс. Проте це досить складна рефлекторна реакція, оскільки одночасно зі скороченням м'язів — згиначів кінцівки відбувається розслаблення її м'язів-розгипачів. В основі такої взаємодії м'язів лежить координація центрів спинного мозку:
збудження центру згиначів через вставні нейрони спричинює одночасне гальмування центру розгиначів Таке гальмування називають реципрокним .
Значення процесів гальмування у функціонуванні нервової системи надзвичайно велике:
воно обмежує неконтрольоване поширення збудження у ЦНС, забезпечуючи чіткі координовані реакції;
відіграє охоронну роль, захищаючи нейрони від ушкодження внаслідок надмірних за силою подразнень.
Основна роль центрального гальмування полягає у взаємодії з центральним збудженням, що полегшує й покращує аналіз і синтез у ЦНС (координаційна роль). Окремі види гальмування відіграють і захисну (оберігальну) роль. Крім того, центральне гальмування здійснює точне дозування сигналів.
Зворотне гальмування (як і слідове) обмежує максимально можливу частоту розрядів мотонейронів спинного мозку і координує (узгоджує) максимальну частоту розрядів із швидкістю скорочення м'язових волокон, які іннервує, та відіграє захисну роль (забезпечує мотонейрони від перезбудження).
_ЗАКЛЮЧЕННЯ.
Підводячи підсумки лекції, можна зробити такі висновки :
1 – для узгодженої роботи нервової систесми в ній постійно повинні відбуватися процеси збудження і гальмування,
2 –збудження дозволяє сприйняти інформацію і обробити її, а гальмування – відсікти зайву інформацію,
3 – процес збудження може розповсюджуватися, гальмування є локальним процесом-+,
4 – гальмівний процес доволі складний і має первні особливості залежно від виду гальмування,
5 - знання особливостей процесів збудження і гальмування нервової системи дозволить більш глибоко зрозуміти особливості нервової регуляції організму, що для медичного працівника має велике значення.
А тепер для того , щоб з`ясувати , наскільки добре ви засвоїли матеріал лекції проведемо невелике тестування .