

Метаболізм кетонових (ацетонових) тіл
Кетоновими (ацетоновими) тілами називають ацетооцтову кислоту (ацетоацетат) СН3-СО-СН2-СООН, -гідрооксимасляну кислоту (-оксибутират) СН3-СНОН-СН2-СООН і ацетон СН3-СО-СН3. Вони утворюються в печінці з ацетил-КоА. На першій стадії цього процесу з двох молекул ацетил-КоА утворюється ацетоацетил-КоА.
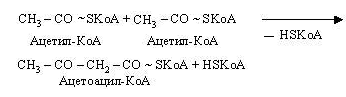
Потім ацетоацетил-КоА взаємодіє ще з одною молекулою ацетил-КоА.
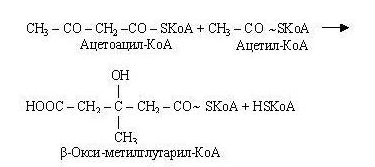
-гідрокси--метилглутарил-КоА, що утворився, розщеплюється на ацетоацетат і ацетил-КоА.
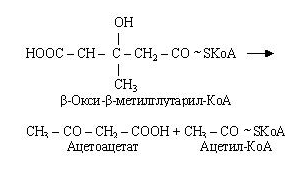
Ацетоацетат (ацетооцтова кислота) при участі НАДН2 відновлюється до -гідромасляної кислоти (-гідрооксибутират).
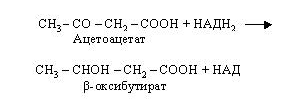
Крім того, ацетоацетат при декарбоксилюванні перетворюється в ацетон. Ацетонові тіла розглядаються як специфічні, а не проміжні продукти обміну біосинтезу жирних кислот.
В крові здорової людини кетонові тіла містяться лише в невеликих кількостях.
При важких формах діабету, повному голодуванні, недостатньому змісті вуглеводів у їжі концентрація кетонових тіл у крові збільшується і може досягати 20 ммоль/л, розвивається ацидоз (кетоз), при цьому швидкість утворення кетонових тіл перевищує здатність периферичних тканин їх утилізувати.
В даний час доведена важлива роль кетонових тіл у підтримці енергетичного гомеостазу. Вони є джерелами енергії для роботи м'язів, нирок і діють як частина регуляторного механізму зі зворотним зв'язком, запобігаючи надзвичайній мобілізації жирних кислот з "жирових депо". Отже, розвиток гіперліпемії сприяє заощадженню вуглеводів при їхньому недоліку. Виключенням є печінка і головний мозок, що не використовують кетонові тіла як джерело енергетичного матеріалу. У м'язах і інших тканинах ацетонові тіла окисляються з утворенням АТФ.
12.3. Біосинтез ліпідів
Здатність людини і тварин запасати глікоген обмежена, тому кількість глюкози, що надходить в організм, може перевищувати його енергетичні потреби. При цьому глюкоза використовується в якості "будівельного матеріалу" для синтезу жирних кислот і гліцеролу, що перетворюються в триацилгліцерол, що відкладається в "жирових депо".
У синтезі триацилгліцеролів умовно виділяють три етапи: утворення гліцеролу, синтез жирних кислот, взаємодія гліцеролу і вищих жирних кислот з одержанням складних ефірів.
Гліцерол у виді гліцерофосфату утвориться або з продукту розпаду вуглеводів 3-фосфо-гліцеролового альдегіду, або з гліцерофосфату, що виділяється при розщепленні триацилгліцеролів у клітках тканин.
Біосинтез жирних кислот полягає в поступовому подовженні ланцюга жирної кислоти на два вуглецевих атоми до виникнення визначеної жирної кислоти. У цьому процесі беруть участь ацетил-КоА, малоніл-КоА, АТФ, НАДФН2, ацилпереносячий білок і система ферментів, з’єднаних у мультиферментний комплекс - синтетаза жирних кислот.
"Стартовою" речовиною біосинтезу жирних кислот є малоніл-КоА. Він утворюється з ацетил-КоА шляхом приєднання активованого оксиду вуглецю.
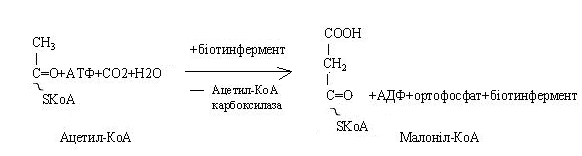
Синтетаза складається із семи ферментів, що утворюють сферичну частку, у центрі якої розташований ацетилпереносячий білок (АПБ). До нього при синтезі приєднуються вихідні субстрати - ацетил-КоА, малоніл-КоА, і він переміщає їх від одного ферменту до іншого.
На початку синтезу відбувається приєднання ацетил-КоА і малоніл-КоА до синтетази при участі АПБ і утворюється ацетоацетил-АПБ. Потім він переміщається до наступного ферменту (редуктази), що передає йому два атоми гідрогену від НАДФ·Н2. Речовина, що утворюється, взаємодіє з ферментами, що входять до складу синтетази. Цим закінчується один цикл синтезу, і карбогеновий ланцюг кислоти подовжується на два карбогенових атоми.
Не відщеплюючись від АПБ, жирна кислота включається в наступний цикл синтезу, що починається з приєднання до неї малоніл-КоА. Цей процес повторюється доти, поки не синтезується потрібна для клітини жирна кислота з визначеною кількістю карбогенових атомів. Кислота, що утворилася, відщеплюється від синтетази, вступає в інші реакції, а до синтетази приєднуються вихідні субстрати, і починається синтез наступної молекули насиченої жирної кислоти.
Ненасичені жирні кислоти в організмі людини не синтезуються, тому вони повинні надходити з їжею.
На останньому етапі синтезу триацилгліцеролів відбувається взаємодія гліцерофосфату з двома молекулами активованих жирних кислот. Спочатку утворюється фосфатидна кислота, що є проміжним продуктом при утворенні триацилгліцеролів і фосфоліпідів.
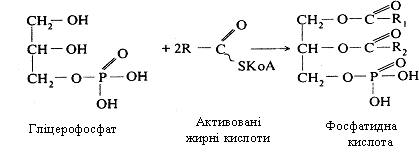
Потім від цієї кислоти відщеплюється фосфорна кислота, до диацилгліцеролу, приєднується ще одна молекула активованої жирної кислоти і утворюється триацилгліцерол (рис. 12.2.).
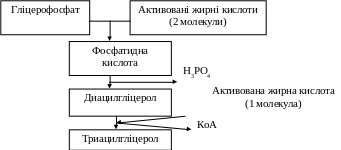
Рис 12.2. Синтез тригліцеролів
У процесі синтезу фосфоліпідів з'являються диацилгліцерол і активна форма холіну - цитідиндифосфохолін (ЦДФ-холін), що взаємодіють між собою з утворенням лецитину. Найбільше активно цей процес відбувається в печінці, з якої вони розносяться до органів і тканин (рис. 12.3.).
Гліцерофосфат
Активовані жирні кислоти (2 молекули)
Холін


АТФ
Фосфатидна кислота
Фосфохолін
H3PO4
ЦТФ
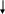

Диацилгліцерол
Цитидін-фосфохолін



Фосфатидилхоліни (лецитини)
Рис. 12.3. Синтез фосфатидилхолінів.