

Аеробне окиснювання вуглеводів
Клітини, які недостатньо постачаються киснем, можуть частково або повністю існувати за рахунок енергії гліколізу.
В аеробних умовах перетворення лактату, що утворився із глюкози в процесі гліколізу, іде у двох напрямках: приблизно 1/5 лактату окисляється до СО2 і Н2О, інша кількість перетворюється в глікоген.
Перший етап аеробного розпаду лактату - його окиснювання до пірувату за участю лактатдегідрогенази.
Однак переважна більшість тваринних і рослинних клітин у нормі перебуває в аеробних умовах, і піруват, що утворюється при розщепленні глюкози, не відновлюється до лактату, а поступово окисляється до СО2, і Н2О в аеробної стадії катаболізму. При цьому спочатку відбувається окисне декарбоксилювання пірувату з утворенням ацетил-КоА.
Окисне декарбоксилювання протікає на мітохондріях. Воно каталізується складними поліферментними і полікоферментними системами, які складаються з декількох ферментів (власне дегідрогенази й ін.) і декількох коферментів, у тому числі тіамінпірофосфату (ТПФ), тіаміндифосфату (ТДФ), ліпоєвої кислоти (ЛК), НАД, HSKoА, ФАД, Mg2+. Кожен компонент ферментної системи бере участь у здійсненні відповідної ланки процесу. Реакції відбуваються в кілька етапів.
Цикл трикарбонових кислот (цикл Кребса)
У ході аеробного окиснювання вуглеводів піровиноградна кислота піддається окисному декарбоксилюванню з утворенням ацетил-КоА.
Ацетил-КоА, що утворився, вступає в різні реакції обміну й окислюється до СО2 і Н2О. Повне окиснювання ацетил-КоА відбувається в циклі трикарбонових кислот - циклі Кребса.
70 % ПВК окисляється до СО2 і Н2О через стадію утворення ацетил-КоА. При обміні білків, жирів і вуглеводів протягом доби на кожен 1 кг маси тіла утворюється близько 10 г ацетату.
Цикл Кребса є центром, де сходяться всі метаболічні шляхи, він - загальний кінцевий шлях окиснювання ацетильних груп (у вигляді ацетил-КоА), у які перетворюється в процесі катаболізму більша частина органічних молекул, що грають роль "клітинного палива" - вуглеводів, жирних кислот, амінокислот.
Він протікає в матриксі мітохондрій і складається з восьми послідовних реакцій.
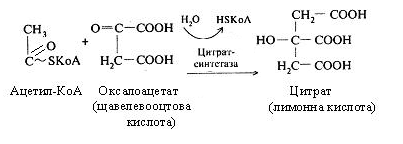
Перша реакція каталізується цитрат-синтетазою, при цьому ацетил-КоА конденсується із щавелевооцтовою кислотою (оксалоацетатом), у результаті чого утворюється лимонна кислота (цитрат).
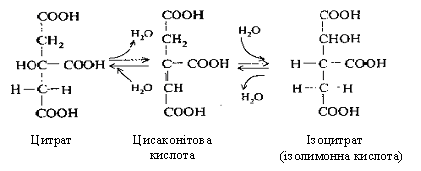
У другій реакції лимонна кислота піддається дегідратуванню з утворенням цисаконітової кислоти, що, приєднуючи молекулу води, переходить в ізолимонну кислоту (ізоцитрат).
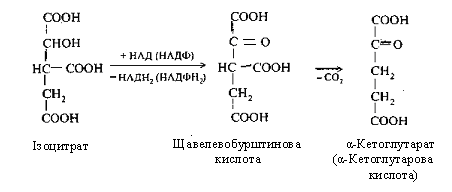
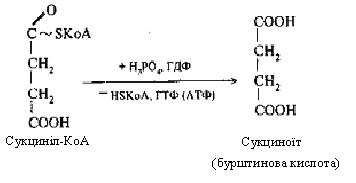
Третя реакція - ізолимонна кислота окиснюється шляхом відщіплення двох атомів водню й перетворюється в щавелевобурштинову кислоту, що декарбоксилюється до α-кетоглутарової кислоти (α-кетаглутарат).
Четверта реакція: α-кетоглутарова кислота піддається окисному декарбоксилюванню під впливом складних ферментів До складу цих ферментів входять коферменти: (ТПФ, HSKoА, ліпоєва кислота, НАД, ФАД). Для реакції необхідні також іони Mg2+. У результаті окисного декарбоксилювання α-кетоглутарової кислоти утворюється сукциніл-КоА, що має макроергічний карбоксилтіоловий зв'язок, у якому резервується енергія окисного декарбоксилювання. При цьому виділяється друга молекула СО2 і НАДН2.
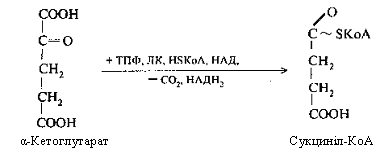
У ході п'ятої реакції сукциніл-КоА взаємодіє з неорганічним фосфатом, потім реагує з гуанідилдифосфатом (ГДФ) і перетворюється в бурштинову кислоту, а ГДФ, приєднуючи фосфат, переходить у ГТФ.
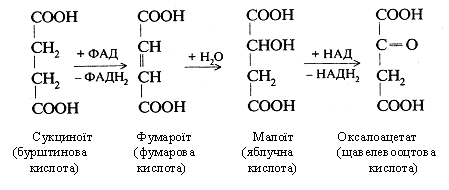
У результаті шостої реакції бурштинова кислота при участі ФАД окисляється у фумарову кислоту (фумарат). Остання гідратується з утворенням яблучної кислоти (сьома реакція). Під впливом НАД вона окисляється до щавлевооцтової кислоти (оксалоацетат) - восьма реакція, тобто до того субстрату, з якого починається цикл трикарбонових кислот.
Процес аеробного окиснювання вуглеводів відбувається з виділенням енергії (за рахунок НАДН2 і ФАДН,). Одна молекула НАДН2 виділяється на стадії декарбоксилювання пірувата з утворенням ацетил-КоА.
Окиснювання ізолимонної кислоти в щавелевобурштинову також супроводжується виділенням молекули НАДН2. При декарбоксилюванні α-кетоглутарової кислоти утворюється сукциніл-КоА й НАДН2. Сукциніл-КоА перетворюється в бурштинову кислоту з утворенням 1 молекули АТФ (за рахунок ГТФ). Дегідрування бурштинової кислоти відбувається з виділенням ФАДН2, а дегідрування яблучної - з виділенням НАДН2. Відомо, що 1 молекула НАДН2 еквівалентна 3 молекулам АТФ, а 1 молекула ФАДН2 - 2 молекулам АТФ.
Отже, з однієї тріози в аеробному циклі утворюється 4 молекули НАДН2 (12 молекул АТФ), 1 молекула ФАДН2 (2 молекули АТФ) і 1 молекула АТФ (ГТФ). Оскільки з однієї молекули глюкози утворюється 2 тріози, вихід енергії буде становити 15 х 2 = 30 молекул АТФ, а повне окиснювання 1 молекули глюкози в анаеробному і аеробному циклах складе 8 + 30 = 38 молекул АТФ.
Щавелевооцтова кислота, що утворилася, може вступати в реакцію з іншою молекулою ацетил-КоА й процес починається спочатку. Таким чином, певна кількість щавелевооцтової кислоти неодноразово бере участь у реакції й забезпечує окиснювання до СО2 і Н2О великої кількості оцтової кислоти, що утворюється з вуглеводів, ліпідів, білків й інших сполук. Але цей процес не нескінченний. Щавелевооцтова кислота в мітохондріях декарбоксилюється з утворенням піровиноградної кислоти, що у свою чергу перетворюється в ацетил-КоА.
Виділенням енергії не вичерпується значення циклу трикарбонових кислот. Роль його полягає також в тому, що при аеробному окиснюванні вуглеводів утворюються проміжні речовини, які використовуються для різних процесів біосинтезу. Так, α-кетоглутарова й щавелевооцтова кислоти можуть перетворюватися в амінокислоти (глутамінову й аспарагінову), необхідні для синтезу білків, азотистих основ, гормонів, нуклеотидів.
Цикл Кребса поставляє відбудовні еквіваленти в ланцюг дихальних ферментів, де потік електронів і протонів сполучений з утворенням АТФ. Цикл не може протікати швидше, ніж це дозволяє використання АТФ. Він регулюється за типом зворотного зв'язку: інгібується АТФ й активується АДФ, що накопичується в міру утилізації АТФ.