

Декарбоксилювання амінокислот
Процес декарбоксилювання каталізується декарбоксилазами, специфічними для кожної амінокислоти, простетичною групою яких служить піридоксальфосфат. Ці ферменти відносяться до класу ліаз. Процес декарбоксилювання, що полягає у відщіпленні від амінокислот СО2 з утворенням амінів, можна показати на наступній схемі:

Механізм реакції декарбоксилювання амінокислот відповідно до загальної теорії піридоксалевого каталізу зводиться до утворення піридоксальфосфат-субстратного комплексу в активному центрі ферменту.
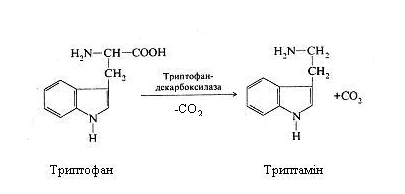
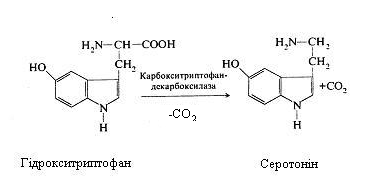
Таким шляхом із триптофану утворюється триптамін, з гідрокситриптофана - серотонін.
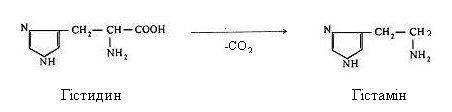
З амінокислоти гістидину утворюється гістамін.
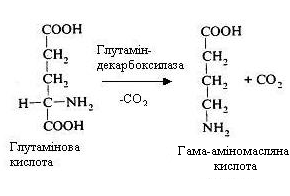
З глутамінової кислоти при декарбоксилюванні утворюється -аміномасляна кислота (ГАМК).
Аміни, утворені з амінокислот, називають біогенними амінами, тому що вони виявляють на організм могутній біологічний ефект.
Біогенні аміни виявляють фізіологічну дію в дуже малих концентраціях. Так, введення в організм гістамину приводить до розширення капілярів і підвищення їхньої проникності, звуження великих судин, скорочення гладких м'язів різних органів і тканин, підвищення секреції соляної кислоти в шлунку. Крім того, гістамін бере участь у передачі нервового порушення.
Серотонін сприяє підвищенню кров'яного тиску і звуженню бронхів; його малі дози придушують активність центральної нервової системи, у великих дозах ця речовина робить стимулюючу дію. У різних тканинах організму великі кількості гістаміну і серотоніну знаходяться в зв'язаній, неактивній формі. Біологічну дію вони виявляють тільки у вільній формі.
Гама-аміномасляна кислота (ГАМК) накопичується в мозковій тканині і являє собою нейрогуморальний інгібітор-медіатор гальмування центральної нервової системи.
Великі концентрації цих сполук можуть являти загрозу для нормального функціонування організму. Однак у тваринних тканинах мається аміноксидаза, що розщеплює аміни до відповідних альдегідів, що потім перетворюються в жирні кислоти і розпадаються до кінцевих продуктів.
10.5. Процеси знешкодження аміаку
У процесі перетворення амінокислот у тканинах утворюються їхні кінцеві продукти обміну - оксид карбогену, вода й аміак. Вода використовується організмом для забезпечення біохімічних процесів. Оксид карбогену частково виводиться з організму з видихуваним повітрям, інша його частина утилізується в процесах синтезу (наприклад, при синтезі жирних кислот, пуринових основ і т.д. ). Аміак, що утворюється в результаті дезамінування амінокислот, є токсичною речовиною, збільшення його концентрації в крові й інших тканинах робить несприятливу дію, особливо на нервову систему. Токсичність аміаку обумовлена тим, що він сприяє відбудовному амінуванню -кетоглутарової кислоти в мітохондріях. Це приводить до видалення її з циклу Кребса і, як наслідок, до падіння тканинного дихання і надлишкового утворення кетонових тіл з ацетил-КоА.
У процесі еволюції живі організми виробили різні ефективні механізми зі знешкодження токсичної дії аміаку, основними з яких є: утворення амінів глутаміну або аспарагіну, відбудовне амінування, нейтралізація кислот, синтез сечовини.
Синтез глютаміну або аспарагіну має велике значення для організму. Він протікає в місцях безпосереднього утворення аміаку (наприклад, у печінці, мозку), там же знаходиться і фермент, каталізуючий цей процес, глутамінсинтетаза, що відноситься до класу лігаз. Синтез амідів вимагає доставки енергії у виді АТФ, присутності глутамінової або аспарагінової кислот, вільного аміаку і відповідних специфічних ферментів. Реакція синтезу аміду пов’язана з розпадом АТФ.
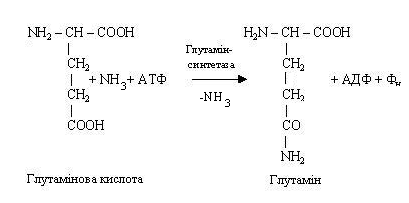
Аналогічно утворюється і аспарагін.
У результаті взаємодії аміаку з глутаміновою і аспарагіновою кислотами відбувається його зв'язування, і в такий спосіб аміак знешкоджується. Зв'язаний аміак може бути використаний як джерело азоту (наприклад, для синтезу пуринових і піримідинових основ, мукополисахаридів). Глутамін і аспарагін не тільки знешкоджують аміак, але і виступають у якості його транспортної форми. У зв'язаному виді аміак доставляється до місця остаточної утилізації - у печінку, де з нього синтезується сечовина.
Синтез сечовини. Одним з найбільш ефективних методів знешкодження аміаку є синтез сечовини. Вперше схема синтезу сечовини була запропонована російським біохіміком М. В. Ненцким, який вважав, що сечовина утворюється з двох молекул аміаку й однієї молекули вугільної кислоти. При цьому у вугільній кислоті гідроксильні групи заміщаються аміаком з утворенням проміжної сполуки карбоновокислого амонію.
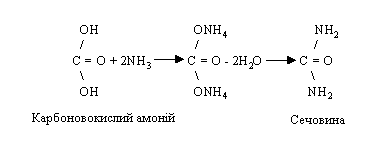
Однак надалі експериментального підтвердження ця теорія не знайшла. Карбоновокислий амоній у печінці практично не виявляється. Крім того, ця речовина не менш отрутна, ніж аміак.
У спеціальних дослідах М. В. Залесского і С. С. Салазкіна на тваринах (лабораторія акад. І. П. Павлова) було встановлено, що якщо венозну кров направити не в печінку, а, минаючи її, у нижню порожню вену, то спостерігаються різке збільшення вмісту аміаку в крові й отруєння. На підставі цих результатів був зроблений висновок, що печінка є органом, у якому відбувається знешкодження аміаку.
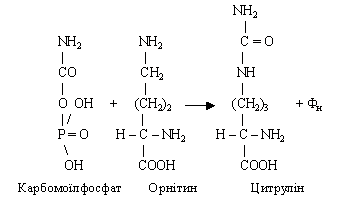
У роботах X. А. Кребса й інших дослідників була показана участь у даному процесі речовин і ферментів, каталізуючих реакції синтезу сечовини. Англійський біохімік Х. Кребс вніс великий вклад у сучасну теорію синтезу сечовини. Він встановив, що цей процес носить циклічний характер, і вказав на роль у ньому орнітину. Передумовою для створення теорії синтезу сечовини було виявлення в печінці амінокислоти орнітину і ферменту аргінази, що розщеплює аргінін на орнітин і сечовину. За теорії Кребса, синтез сечовини починається взаємодією орнітину з аміаком і оксидом карбогену. При цьому утворюється цитрулін, що взаємодіє з ще одною молекулою аміаку з виділенням аргініну. Останній гідролізується аргіназою на орнітин і сечовину. Орнітин у цих реакціях виконує роль каталізатора. За відкриття цього циклу X. А. Кребс був визнаний гідним Нобелівської премії.
Замкнутий цикл синтезу сечовини умовно можна розділити на три етапи. На першому і другому етапах відбувається зв'язування двох молекул аміаку в нешкідливу для організму сполуку, на третьому етапі утворюється сечовина.
Перший етап. За рахунок енергії АТФ із молекули аміаку й оксиду карбогену синтезується карбомоїлфосфат. Потім відбувається конденсація карбомоїлфосфату з орнітином, у результаті якої синтезується цитрулін. При цьому вивільняється неорганічний фосфат.
Синтез цитруліну забезпечується за рахунок енергії, акумульованої в карбомоїлфосфаті.
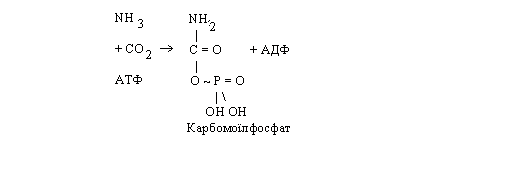
Таким чином, у цитруліні зафіксована одна молекула аміаку.
Другий етап. Відбувається зв'язування ще однієї молекули аміаку в глутамінову кислоту шляхом відбудовного амінування з -кетоглутаровою кислотою. Глутамінова кислота передає зафіксовану молекулу аміаку у виді NH2-групи на щавелевооцтову кислоту, що перетворюється в аспарагінову кислоту (процес переамінування):
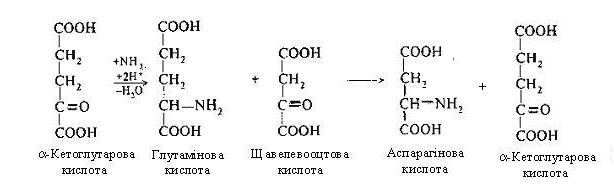
Третій етап. На цьому етапі здійснюється синтез сечовини. Цитрулін, взаємодіючи з аміногрупою аспарагінової кислоти, утворює аргінінобурштинову кислоту (аргінінсукцинат).
У цьому процесі використовується енергія ще однієї молекули АТФ.
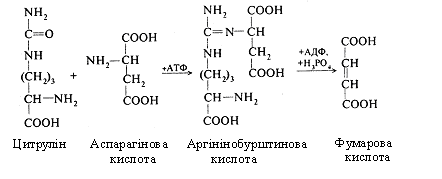
Аргінінобурштинова кислота розщеплюється на аргінін і фумарову кислоту. Фумарова кислота включається у вуглеводний обмін, перетворюючись в яблучну, котра шляхом дегідрування переходить у щавелевоцтову.
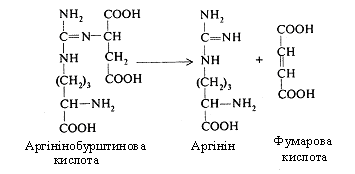
Щавелевооцтова кислота може приєднувати до себе аміак і знову перетворюватися в аспарагінову кислоту або, піддаючись декарбоксилюванню, у піровиноградну, а потім у коензим А, що використовується в різних реакціях біосинтезу або окисляється до СО2 і Н2О.
Аргінін, що утворився, під дією аргінази гідролітично розщеплюється на орнітин і сечовину.
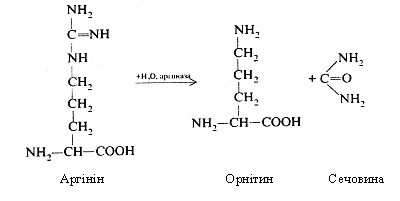
Орнітин, що вивільнився, може знову вступити в реакцію з новою молекулою карбомоїлфосфату і процес повториться.
Підраховано, що в стані азотистої рівноваги організм дорослої людини споживає і відповідно виділяє 15 г азоту; з екскретованого із сечею азоту на долю сечовини приходиться близько 85 %, креатініну - 5, амонійних солей - 3, сечової кислоти - 1, інших його форм - 6.