

Типы деления клеток
Митоз - тип деления клеток, при котором из одной диплоидной клетки (2п=46) образуются две диплоидные, генетически равнозначные клетки. Процесс митоза в соматических клетках человека идет стандартно. К концу профазы хромосомы становятся отчетливо видимыми, каждая состоит из двух хроматид. Обе сестринские хроматиды прилежат одна к другой. Центромера в каждой хромосоме удерживает две сестринские хроматиды вместе. Ядрышко исчезает, ядерная оболочка распадается на фрагменты. Хромосомы располагаются в цитоплазме в центральной части клетки, оттесняя все органоиды к периферии.
Во время метафазы центромеры всех хромосом располагаются в экваториальной плоскости между двумя полюсами. Хроматиды каждой хромосомы начинают отделяться одна от другой, оставаясь соединенными только в центромерной области. В районе центро-мер с противоположных сторон прикреплены нити веретена деления. Их количество может достигать нескольких десятков в районе каждой центромеры.
Анафаза начинается с одновременного разделения всех центромер и расхождения сестринских хроматид каждой хромосомы к противоположным полюсам. Утрата синхронности процесса может привести к неправильному расхождению хромосом. Центромеры с помощью нитей веретена деления увлекают за собой дочерние хроматиды к противоположным полюсам. Анафаза заканчивается с прекращением движения хроматид, которые становятся хромосомами. У каждого полюса клетки должно оказаться по 46 хромосом, состоящих из одной хроматиды.
Телофаза связана с образованием ядерных оболочек вокруг хромосом на двух полюсах клетки и началом перехода хромосом в состояние хроматина. Завершается телофаза образованием перетяжки в центральной части делящейся клетки, которая завершает деление клетки надвое.
Известно, что у эукариот прохождение митоза может быть заблокировано физиологически или экспериментально, что приводит к развитию полиплоидных клеток. Системная полиплоидия не характерна для человека. Полиплоидные эмбрионы погибают на ранних стадиях развития, что является одной из причин спонтанного прерывания беременности на ранних сроках. В то же время нали чие полиплоидных клеток в некоторых органах человека не является патологией. Например, полиплоидные клетки встречаются в сердечной мышце, особенно в предсердиях, в печени и некоторых железах. Они могут быть одно- и двуядерными, уровень полиплоидии невысок и обычно находится в пределах 4—8п.
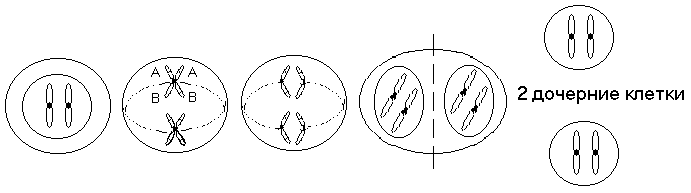
Мейоз - особый тип клеточного деления, возникновение которого связано с появлением полового размножения. При половом размножении два родителя — отец и мать — дают начало новому организму. При оплодотворении сливаются ядра половых клеток родителей, что увеличивает вдвое количество хромосом в зиготе. Следовательно, образование половых клеток должно быть связано с уменьшением количества хромосом в два раза, но таким образом, чтобы совокупность генетического материала
обеспечивала преемственность поколений. Закономерное чередование репликации ДНК и, соответственно, xromocom, митозов и мейозов обеспечивает сохранение видоспецифического кариоти-па как в индивидуальном развитии — онтогенезе, так и в череде поколений организмов.
В процессе мейоза из одной диплоидной клетки (2п=46) образуются 4 гаплоидные клетки (п=23). Кроме того, в мейозе происходит два вида перегруппировки генетического материала хромосом, т. е. два вида генетической рекомбинации: 1) независимое распределение гомологичных хромосом из разных пар к полюсам деления; 2) кроссинговер — обмен участками между гомологичными хромосомами. Эти процессы обеспечивают широчайший спектр наследственной изменчивости, генетическую неповторимость индивидов даже среди потомков одной пары родителей.
Мейотическое деление у человека не имеет каких-то коренных отличий от мейоза других эукариот. Оно состоит из двух, следующих друг за другом делений, между которыми не происходит удвоения ДНК, а следовательно, и хромосом.
Перед мейозом обязательно проходит интерфаза, в S-периоде которой ДНК реплицируется. Следовательно, в профазе первого мейотического деления выявляющиеся нитевидные хромосомы состоят из двух хроматид. Каждое из двух делений мейоза состоит из про-, мета-, ана- и телофазы с индексами I или II.
Первое мейотическое деление протекает значительно дольше, чем второе. Самой длительной фазой первого мейотического деления является профаза, так как именно в этой фазе происходят такие сложные процессы, как образование бивалентов из гомологичных хромосом и кроссинговер.
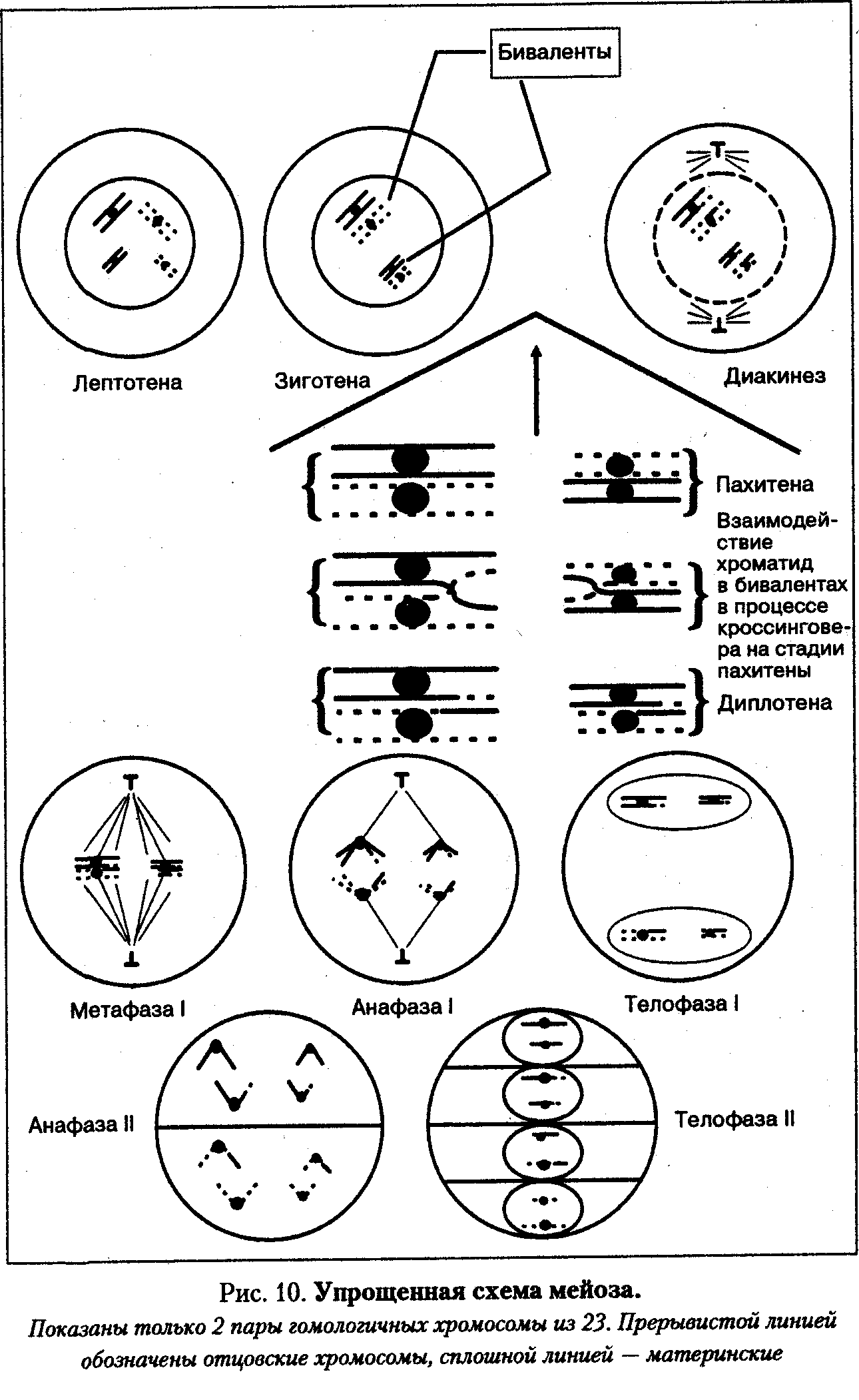
Профаза мейоза I подразделяется на 5 подфаз: лептотена, зиго-тена, пахитена, диплотена, диакинез.
Лептотена — стадия тонких нитей. Ядра клеток, вступающих в мейоз, значительно крупнее других. В этих ядрах вместо хроматина выявляются очень тонкие и длинные нитевидные хромосомы. Они настолько тонкие и длинные, что их тру дно проследить по всей длине. Для лептотены характерно также появление на тонких хромосомах особых, напоминающих бусины структур — хромомеров. Это участки более сильно конденсированного хроматина. Число, размер и расположение хромомерных участков специфичны для каждой хромосомы. Выявление хромомерных структур отражает постепенный процесс конденсации хромосом из хроматина. Каждая хромосома состоит из двух сестринских хроматид, но их далеко не всегда удается различить под световым микроскопом, настолько близко они прилегают друг к другу.
Зиготена - стадия сливающихся нитей. На этой стадии гомологичные хромосомы находят друг друга и сливаются (конъюгируют) с образованием бивалента. У человека 46 хромосом (23 пары гомологичных хромосом), следовательно, количество бивалентов равно 23 и соответствует гаплоидному набору хромосом.
Бивалент - это стабильная структура, состоящая из двух гомологичных хромосом, или 4-х хроматид. Стабильность этой структуры поддерживается с помощью специфических белков синалто-немного комплекса. Объединение гомологов чаще всего начинается на концах хромосом (в теломерах), а также в центромерных районах. Позднее по длине соединяющихся хромосом (внутри бивалента) между хромосомами формируются белковые тяжи синап-тонемного комплекса, сближающие хромосомы.
В настоящее время показана специфичность хромомерного строения индивидуальных бивалентов человека в мужском и женском мейозе, т.е. по рисунку хромомер можно определить, какая пара хромосом образует тот или иной бивалент, даже если по размеру и общей морфологии биваленты одинаковы. Конъюгация гомологичных хромосом с образованием бивалентов является обязательным процессом для всех хромосом человека, включая короткие и половые хромосомы. Показано, что конъюгация происходит не только между половыми хромосомами Х и X, но также между Х- и Y-хромосомами, несмотря на большую разницу в их морфологии (размерах).
В процессе сперматогенеза половой бивалент из Х- и Y-хромосом начинает конъюгировать раньше других хромосом. Конъюгируют между собой часть района короткого плеча Х- и короткое плечо Y-хромосомы. Эксперименты по гибридизации ДНК показали, что эти районы гомологичны между собой. Негомологичные участки хромосом Х и Y остаются свободными.
Зиготена заканчивается образованием 23 бивалентов. Пахитена — стадия прохождения кроссинговера. В пахитене хромосомы выявляются в виде толстых нитей, так как они представлены бивалентами. Именно в бивалентах происходит кроссинговер — взаимный обмен идентичными участками по длине гомологичных хромосом. Генетическим следствием кроссинговера является рекомбинация сцепленных генов, что обеспечивает широкую генетическую изменчивость гамет. Морфологически этот процесс в пахитене уловить нельзя. Для умозрительного восприятия его можно изобразить на схеме (пахитена и диплотена). Кроссинговер — это процесс, происходящий со сложными пространственными изменениями суперспирализованных участков молекул ДНК несестринских хроматид с использованием целого комплекса ферментов, возможно объединенных вместе в специализированную структуру. Кроссинговер является обязательным процессом в каждом биваленте. Не исключено, что отсутствие кроссинговера в каком-то биваленте может быть запирающим фактором мейоза.
В дальнейшем, в диплотене, когда начинают расходиться гомологичные хромосомы, в бивалентах, выявляются те места, где происходил процесс кроссинговера. Это места длительного соединения гомологичных хромосом во время разрушения бивалентов. Поскольку морфологически они напоминают греческую букву «X», их называют хиазмами. В зоне хиазм видно, что в перекрест вовлекаются только две хроматиды из четырех (по одной от каждого гомолога).
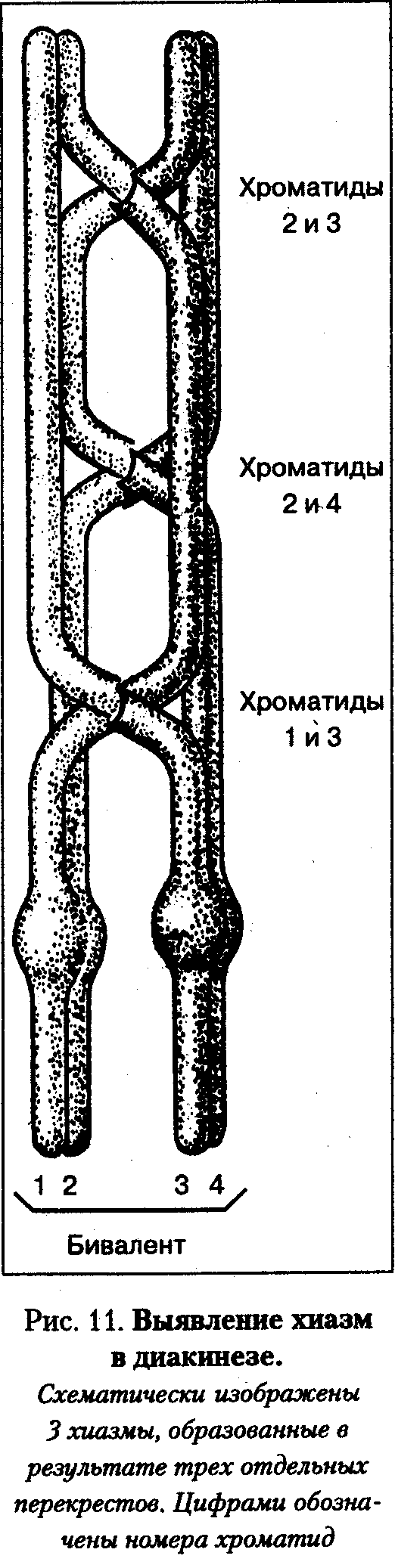
Диплотена – стадия двойных нитей. На этой стадии синаптонемный комплекс разрушается, и гомологи отталкиваются друг от друга, оставаясь соединенными только в районе хиазм, где по-прежнему сохраняется структура синаптонемного комплекса. Так как количество хиазм соответствует одному событию кроссинговера, в котором участвуют две несестринские хроматиды, то по количеству хиазм можно судить об интенсивности процесса кроссинговера. Общее число хиазм на хромосомный набор человека колеблется от 35 до 66. Некоторые биваленты могут содержать несколько хиазм. Среднее количество хиазм на бивалент около двух.
Диакинез — стадия, завершающая профазу мейоза I. Она является переходной к метафазе. Число хиазм уменьшается, биваленты укорачиваются, разрушается ядро, начинает формироваться веретено деления.
Метафаза I — биваленты выстраиваются в экваториальной плоскости в цитоплазме. Центромеры хромосом расположены на экваторе, к ним прикреплены нити веретена деления. Число выстроенных бивалентов соответствует гаплоидному набору хромосом и для человека равно 23.
Анафаза I — расхождение гомологичных хромосом к противоположным полюсам клетки. Каждая хромосома состоит из двух сестринских хроматид.
Телофаза I. В этой фазе происходит образование двух дочерних ядер, каждое из которых содержит гаплоидное число хромосом, равное 23. Каждая хромосома состоит из двух сестринских хроматид.
Промежуток между двумя последующими делениями мейоза очень небольшой. Почти сразу начинается второе мейотическое деление. Оно идет по схеме митоза: 23 хромосомы, состоящие из парных сестринских хроматид, связанных в центромерных участках, в каждом из двух образованных ядер проходят профазу и мета-фазу. В анафазе они разъединяются, и сестринские хроматиды каждой хромосомы расходятся к противоположным полюсам, в результате чего образуется четыре гаплоидных ядра.
Рекомбинация генетического материала в мейозе происходит не только за счет процесса кроссинговера. В анафазе первого мейоти-ческого деления происходит случайное распределение по отношению к полюсам клетки гомологичных хромосом из каждого бивалента. Это приводит к большому числу возможных комбинаций отцовских и материнских хромосом в гаметах. Рассмотрим процесс подробнее на простом примере.
Проанализируем распределение первой и второй пары гомологичных хромосом в анафазе I. Известно, что в каждой паре гомологичных хромосом в процессе оплодотворения одна хромосома приходит из гаметы отца, другая - из гаметы матери. Обозначим заглавными буквами хромосомы отца, а строчными - хромосомы матери. Таким образом, А и а — первая пара хромосом, В и b - вторая пара. В профазе I образуются биваленты. В метафазе I они выстраиваются в экваториальной плоскости: А//а, В// b. В анафазе I гомологичные хромосомы из бивалентов расходятся к противоположным полюсам: к одному полюсу пойдут хромосомы А и В, т.е. отцовские, а к другому, а и b, т.е. материнские. Но у этого события может быть и другой исход, когда расположение хромосом в метафазе на экваторе будет другим: А//а, b//В. Тогда к одному полюсу пойдут хромосомы А и b, а к другому - а и В, т.е. сочетание хромосом на полюсах будет содержать одну отцовскую и одну материнскую. Наличие двух пар гомологичных хромосом обеспечивает, как мы видим, образование четырех типов гамет, качественно отличающихся друг от друга сочетанием отцовских и материнских хромосом. У человека 23 пары хромосом. Разнообразие гамет оценивается как 223. Это примерно 10 миллионов вариантов разнообразных сочетаний отцовских и материнских хромосом из каждой пары гомологов. При оплодотворении практически равновероятна встреча любого из сперматозоидов с овулировавшей яйцеклеткой. Это увеличивает число возможных генотипов детей (223). Частота генетической рекомбинации в результате независимого распределения разных пар гомологов выше, чем частота рекомбинации в результате кроссинговера.