

Спокій Збудження
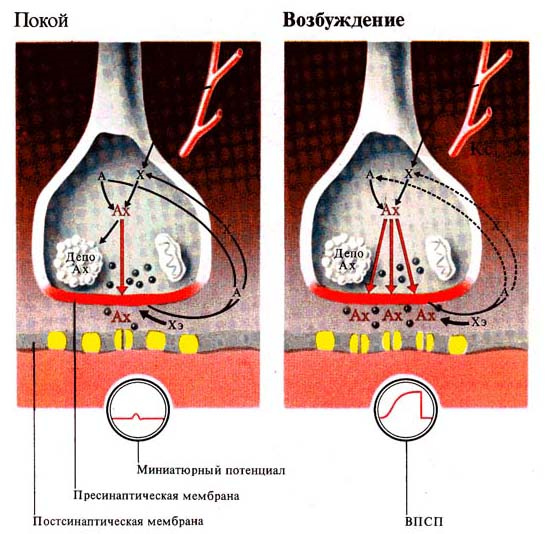
Синаптичні процеси у незбудженому і збудженому синапсах (по л. Щельцину, 1980):
А — ацетат, Х — холін, Хе — холинестераза
Механізм скорочення і розслаблення м'язів
Під час передачі нервового імпульсу через синапс на поверхні мембрани м'язового волокна виникає ПД.
Деполяризація, яка поширюється мембраною, захоплює і мембрани Т-трубочок та цистерн СР, що тісно прилягають до них. Це підвищує проникність мембран СР до іонів кальцію, які за градієнтом концентрації надходять усередину міофібрил і розподіляються між актиновими та міозиновими нитками. Йони кальцію є ініціаторами м`язового скорочення.
Тут іони кальцію виконують дві функції:
По-перше, вони зв'язуються з кальмодуліном, чим відкривають активний центр актину, з яким тут же вступає в реакцію міозиновий місток;
по-друге, нейтралізують негативні заряди АТФ і АТФ-ази містка, внаслідок чого ензим наближається до АТФ і розщеплює її.
Енергія, що виділяється при цьому, витрачається на поворот містка на 45 градусів, який уже зв'язаний з актином.
Після повороту міозинового містка він « випрямляється» і від'єднується від актину, а утворені під час розпаду АТФ неорганічний фосфат і АДФ
відходять.
На їх місце приєднується нова молекула АТФ, що призводить до розриву зв'язку міозину з активним центром актину, - відбувається ковзання ниток актину і міозину одна відносно іншої (на "один крок", або « гребок» рівний 20 нм. Все це нагадує роботу гребця в човні). Водночас інший місток приєднується до наступного актинового центру і т.д.
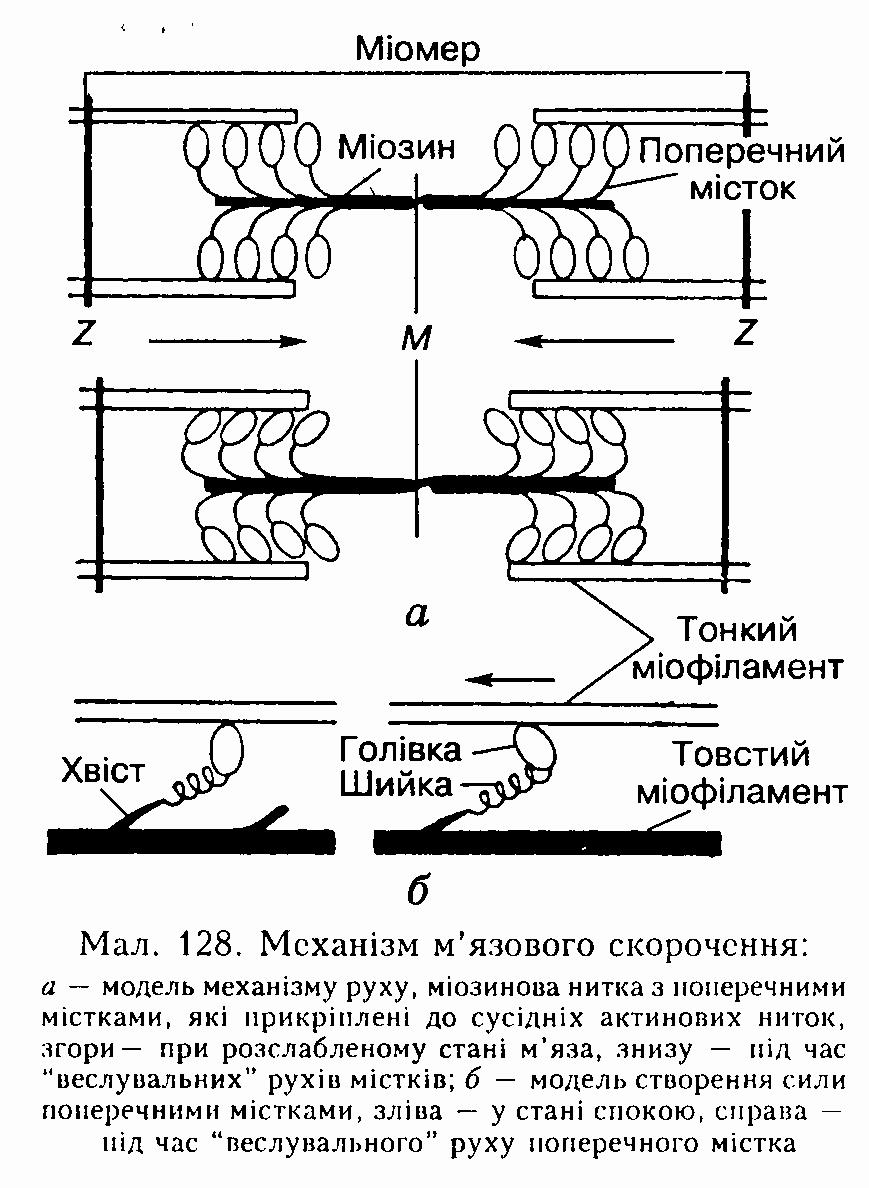
У результаті скорочується кожен саркомер окремо і все м'язове волокно в цілому.
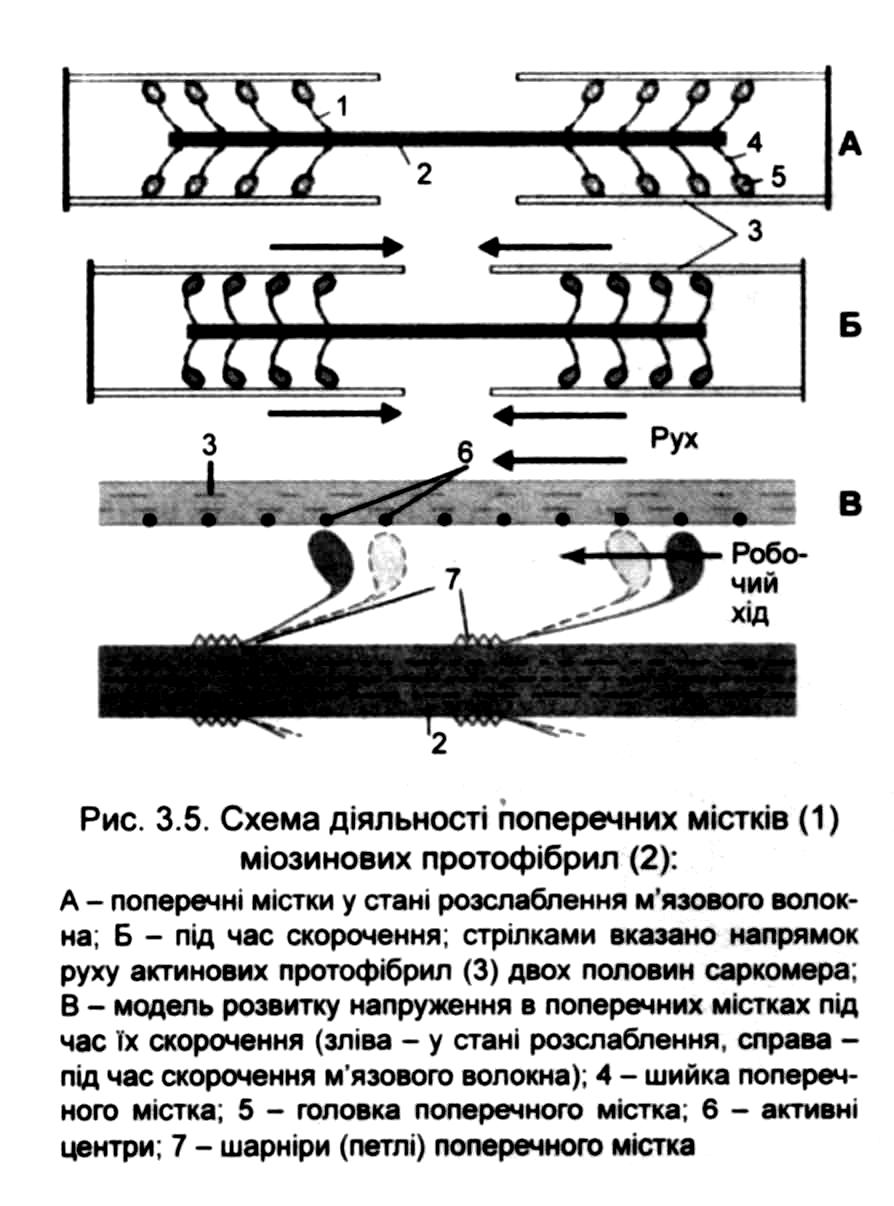
Підсумуємо процеси, що відбуваються в м 'язі при його скороченні:
а) актинові і міозитові нитки практично не коротшають;
б) взаємодія актину з міозином призводить до взаємного входження
ниток у проміжки між ними;
в) в результаті дві сусідні 2-мембрани зближуються одна з одною;
г) оскільки саркоплазма, що міститься всередині саркомера, не
піддається стискуванню, то при зменшенні довжини саркомер розширюється;
д) подібні процеси одночасно протікають у всіх саркомерах м'язового
волокна, тому обидва кінці м'яза підтягуються до центру.
Для розслаблення м'яза необхідно, щоб концентрація іонів кальцію у міофібрилі знизилася.
Це досягається за рахунок роботи кальцієвої помпи на мембранах СР, яка активно перекачує туди іони кальцію.
Актоміозинові зв'язки розриваються і м'яз завдяки своїй пружності та еластичності видовжується.
Оскільки робота помпи вимагає затрат енергії, очевидно, що процес м'язового розслаблення супроводжується розпадом АТФ.
Таким чином, енергія АТФ під час м'язового скорочення і розслаблення витрачається на роботу Nа-К-помпи на поверхні мембрани м'язового волокна (створення мембранного потенціалу), на забезпечення роботи кальцієвої помпи на мембранах СР (створення надлишку іонів кальцію у порожнинах СР) і на саме м'язове скорочення (поворот міозинового містка і ковзання протофібрил).

Схема ковзання ниток актина і міозина в процесі скорочення (по Дж. Бендоллу, 1970):
/ — расслаблення міофибрила, // — розтягнена міофибрила, /// — скорочена міофибрила, зліва — поздовжнє розташування ниток, справа — поперечний переріз.
Електромеханічний зв'язок у м'язі.
При подразненні м'яза через 1-2 мс після піка ПД сила скорочення починає стрімко зростати і скоротлива система повністю активується протягом кількох мілісекунд.
Цей швидкий електромеханічний зв'язок (спряження) між змінами МП і скороченням внутрішньоклітинних міофібрил не може бути зумовлений простою дифузією йонів або медіатору від клітинної мембрани до міофібрил, оскільки така дифузія потребує значно більше часу, ніж 1-2 мс.
Отже, деполяризація клітинної мембрани активує скоротливі елементи якимось іншим чином.
Виявляється, що важливою проміжною ланкою між деполяризацією м'язової мембрани і початком скорочення є проникнення вільних йонів кальцію між міофіламентами.
У стані спокою основна частина Са2+ у м'язовому волокні зберігається в цистернах ендоплазматичної сітки, оскільки проникність мембрани цистерн для Са2+ у спокої мала, а його витік під час скорочення компенсується сталою роботою кальцієвого насоса, який затягує їх назад у цистерни.
Функцію кальцієвого насоса виконує кальційзалежна АТФ-аза (Са2+ АТФ-аза), що розщеплює АТФ із виділенням енергії, яка використовується для закачування Са2+ до цистерн ендоплазматичної сітки. Ось чому концентрація Са2+ у саркоплазматичному матриксу дуже низька (10"8моль/л) у стані спокою волокна і дуже висока (10~5 моль/л) — в цистернах саркоплазматичної сітки.
Вихід Са2+ за концентраційним градієнтом починається під час деполяризації мембрани цистерн ендоплазматичної сітки, що викликає відкриття в ній кальцієвих каналів. Збільшення концентрації Са2+ у саркоплазматичному матриксі відразу після стимуляції було виявлено за допомогою введення у нього білка екворину, який світиться за наявності Са2+.
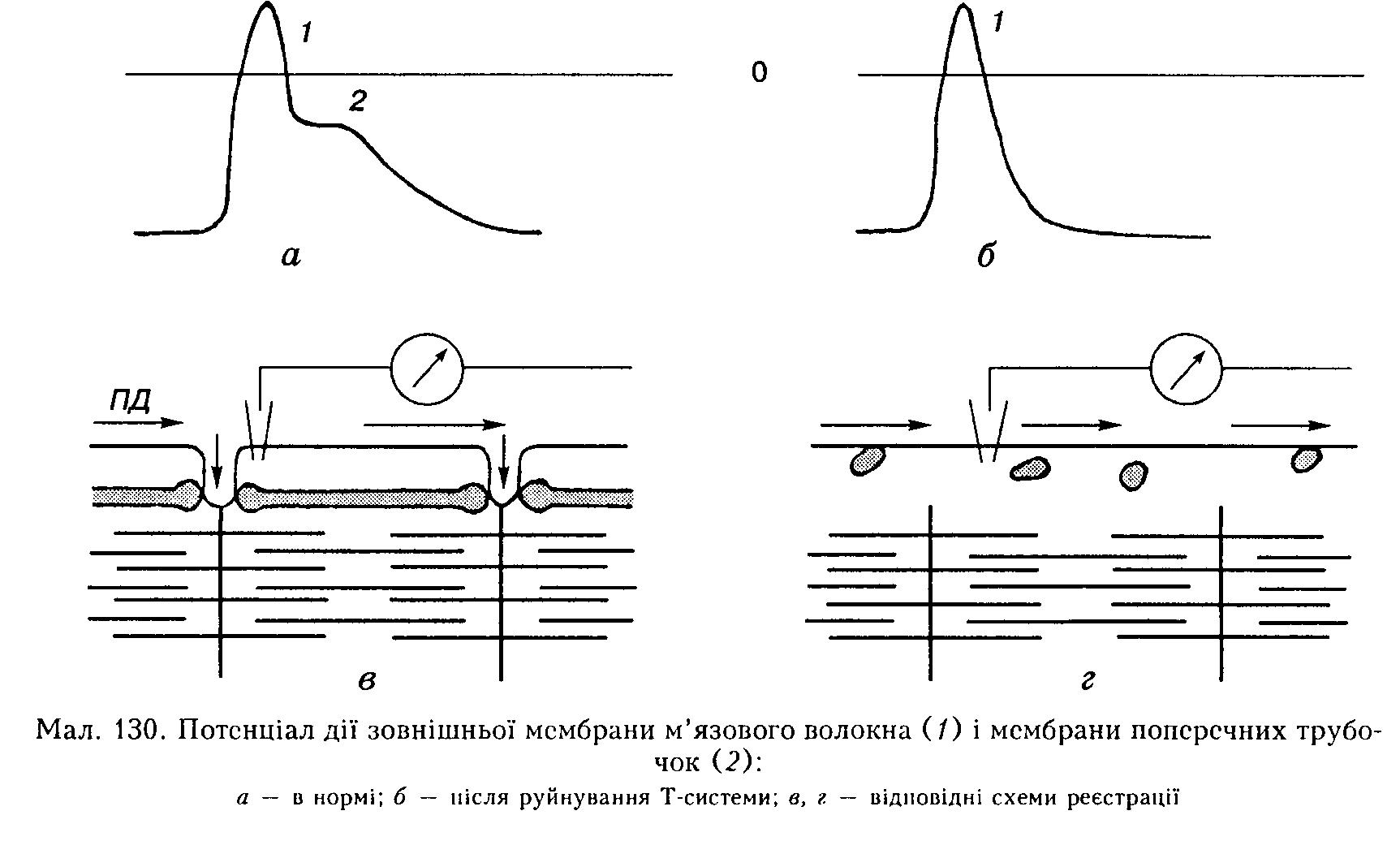
Активація (деполяризація) мембрани цистерн ендоплазматичної сітки відбувається під час поширення ПД сарколеми, зовнішньої м'язової мембрани, на поперечні трубочки Т-системи.
ПД Т-системи м'язового волокна, що виникає при цьому, реєструється внутрішньоклітинним мікроелектродом як додатковий і розтягнутий за часом пік, розміщений па низхідній частині ПД сарколеми (мал. 130, а). ПД поперечної трубочки впливає своїми локальними коловими струмами на мембрану ендоплазматичної сітки, спричинюючи її деполяризацію.
Деполяризація спричинює виділення Са2+ з термінальних цистерн, звідки ці йони одразу потрапляють до тієї ділянки міофібрили, де починається процес скорочення.
Отже, скоротливий акт виникає внаслідок ланцюга послідовних процесів: ПД сарколеми —> ПД поперечних трубочок —> деполяризація мембрани цистерн ендоплазматичної сітки —> вихід Са2+ у саркоплазма-тичний матрикс —> взаємодія Са2+ з міофібрилами (троионін-троиоміозиновою системою) —> скорочення м'язового волокна.
Функціональний електромеханічний зв'язок між сарколемою і міофібрилами може бути порушений штучно, наприклад при обробці м'яза гіпертонічними розчинами гліцерину (400-800 ммоль/л). У цьому випадку Т-система руйнується внаслідок вакуолізації, зникає ПД поперечних трубочок і, хоча є ПД сарколеми, скоротлива реакція не виникає (див. мал. 130, б).
Вивільнення Са2+з цистерн саркоплазматичиої сітки припиняється після закінчення піка ПД, але скорочення триває доти, доки в саркоплазматичному матриксі зберігається висока концентрація Са2+. Лише після її зниження внаслідок роботи кальцієвого насоса поступово настає розслаблення м'язового волокна.
Молекулярний механізм скорочення м'яза.
Насамперед ті йони кальцію, які потрапили з цистерн ендоплазматичної сітки до саркоплазматичного матриксу, тобто у міофібрилярний простір, зв'язуються з тропопіном тонких актинових міофіламентів (два йони кальцію приєднуються до однієї молекули трононіну). Внаслідок цього виникають конформаційні зміни молекули трононіну: вона деформується так, що штовхає тропоміозин у жолобки між закрученими нитками актину — в активоване положення (див. мал. 131, б). При цьому зникає перешкода контакту між голівками молекул міозину товстого міофіламента і мономерами актину тонкого міофіламента, які, з'єднуючись між собою через поперечні містки, утворюють актоміозиновий комплекс (мал. 131, а, б).
Одночасно актоміозиновий комплекс набуває АТФ-азної активності і розщеплює приєднану до голівки молекули міозину молекулу АТФ до АДФ. Відразу після цього голівка молекули міозину за рахунок енергії, виділеної під час розщеплення молекули АТФ, повертається на 45° (див. мал. 128, б), розвиваючи певне напруження, яке передається на її шийку.
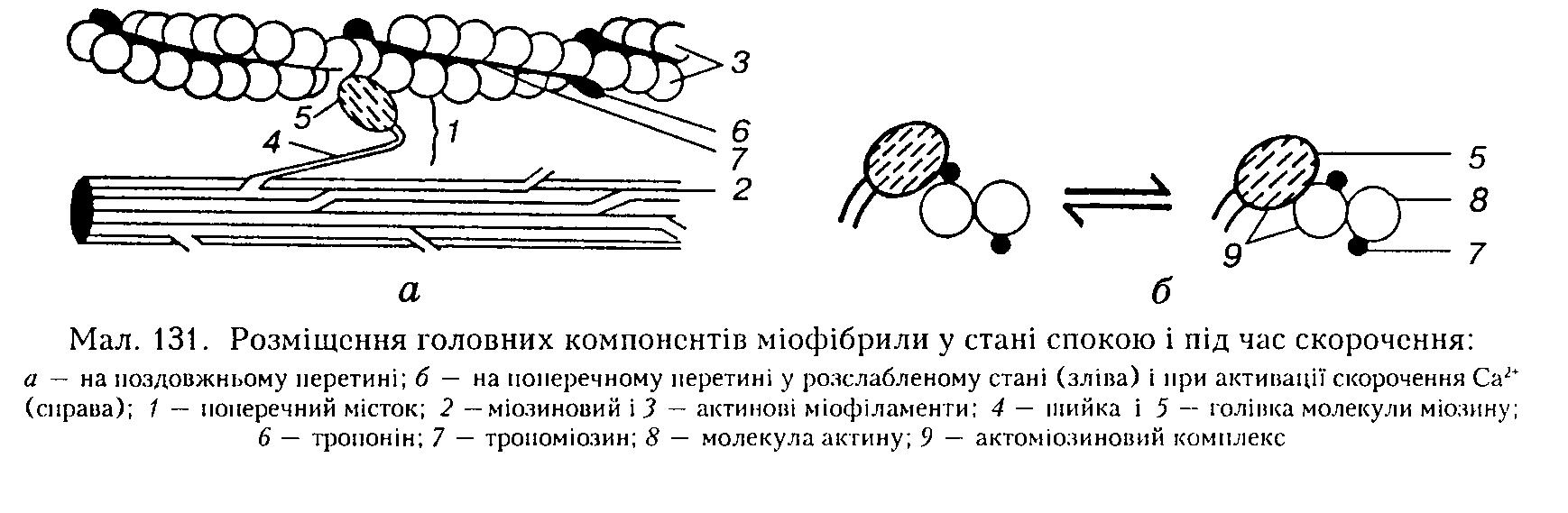
Завдяки еластичним властивостям шийки міофіламенти просуваються (ковзають) один повз одного до центра міомера. Такий поворот голівки молекули міозину нагадує рух весла (див. мал. 128).

За одне таке "веслування" міомер скорочується лише на 20 нм (= 1 % своєї довжини). Після закінчення одного акту "веслування" нова молекула АТФ заміщує молекулу АДФ па голівці молекули міозину, розриває актоміозиновий комплекс, і м'язове волокно розслаблюється. Якщо м'яз скорочується на половину своєї довжини, то в кожному міомері має відбутися 50 веслувальних рухів, кожен з яких ініціюється окремим ПД. У випадках, коли у м'язовому волокні АТФ немає, молекули актину і міозину не роз'єднуються, і утворюється стійкий ригор-комплекс, який виявляється у вигляді заклякання м'язів.
Розщеплення АТФ триває кілька мілісекупд. На цьому цикл перетворення хімічної енергії па механічну закінчується і починається підготовка до другого циклу.
Цей цикл полягає у приєднанні нової молекули АТФ до голівки молекули міозину, роз'єднанні актоміозину, відновленні блокувальної дії тропоміозином молекули актину внаслідок зменшення концентрації Са2+ у саркоплазматичному матриксі, спричиненого закриттям кальцієвих каналів цистерн та постійної роботи кальцієвого насоса.
Отже, під час скорочення м'яза енергія АТФ використовується для трьох головних процесів:
- роботи натрій-калієвого насоса сарколеми під час збудження;
- конформації міозинових молекул та ковзання міофіламентів;
- роботи Са2+-насоса цистерн агранулярної ендоплазматичної сітки.