

Для них характерна пластичність (здатність зберігати розтягнену форму без змін напруження).
Вони утворюють функціональний синцитій завдяки нексусам (міжклітинним контактам без злиття тіл гладеньком'язових клітин - десмосом) і з'єднанням проміжного характеру на зразок відростків.
Мають пейсмекери, тому здатні до автоматії.
Щілинні контакти мають низький опір і забезпечують електротонічне передавання деполяризації від збуджених клітин до сусідніх незбуджених.
Нексуси ( контакти) суміжних клітин містять утворення, подібні до іонних каналів. Відростки відіграють роль у передаванні механічної сили між контактами. Наявність між міоцитами контактів (нексусів) з низьким перехідним опором перетворює їх на електричний синцитій. У результаті багато клітин функціонують як безперервна в електричному відношенні одна велика клітина.
Величина потенціалу спокою цих м'язів коливається від -50 до -60 мВ. Особливості скорочення непосмугованих м'язів
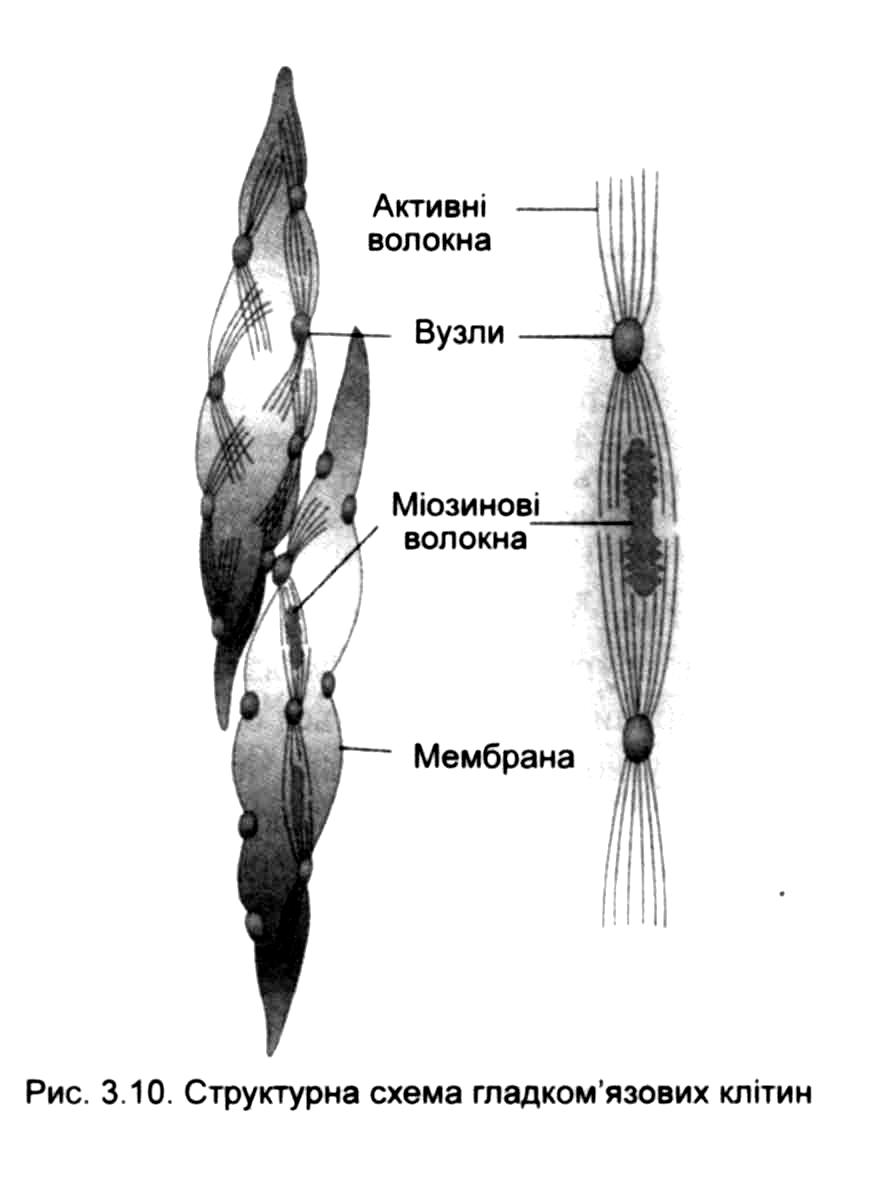
У цитоплазмі непосмугованом'язових клітин наявні актинові та міозинові філаменти, які структурно і функціонально мало відрізняються від відповідних утворів скелетних м'язів, проте розташовані вони не так упорядковано.
Крім того, є і інші особливості :
у непосмугованих м'язах менше міозинових волокон,
немає типових 2-мембран,
у клітині непосмугованих м'язів слабше виражений саркоплазматичний ретикулум.
Актинові філаменти згруповані у пучки, які періодично утворюють ущільнення - вузли. Деякі з них безпосередньо прилягають до мембрани, інші знаходяться всередині клітини, виконуючи функцію 2-мембран. Між актиновими філаментами вклинюються щільніші - міозинові.
Крім актину й міозину вони містять тропоміозин, кальдесмон та кальпонін поздовжнього напрямку. Вони розміщуються під кутом до осі клітини, утворюючи об'ємну сітку.
Міозинові протофібрили гладких м'язів відрізняються від скелетних меншою молекулярною масою, амінокислотним складом, розчинністю, чутливістю до ферментів, солей і денатурації.
Актинові нитки складаються з актину, тропоміозину і кальдесмону. Тропоміозину в гладеньких м'язах менше, ніж у скелетних, і він відрізняється за складом. Актинові нитки містять додаткові білки. Це філамін і вінкулін.
Ділянки двох сусідніх клітин, які прилягають до мембрани, часто збігаються з утворенням тут білкової перемички, що об'єднує структури цих клітин у синцитій.
Скорочення непосмугованих м'язів відрізняється високою економністю порівняно зі скелетними м'язами (АТФ витрачається у 10-100 разів менше), що забезпечує можливість тривалих скорочень без розвитку втоми.
Скорочення гладеньких м'язів - тривалі й менш втомливі.
Умовно скорочення поділяються на фазні (фазно-тонічні) й тонічні.
Фазні генерують повноцінний потенціал дії,
тонічні - лише місцеве збудження, хоча за певних умов також можуть генерувати потенціал дії.
Скоротлива активність гладких м'язів.
Кожний ПД викликає поодиноке скорочення гладкої м'язової клітини. Електромеханічний зв'язок (він називається спряженим) у гладких м'язах здійснюється за допомогою Са2+, які надходять до клітини ззовні, а також виходять із внутрішньоклітинних депо (поперечних трубочок ендоплазматичної сітки). Проте ця сітка в гладких м'язах розвинута недостатньо, тому головну роль у електромеханічному спряженні їх відіграє зовнішньоклітинний кальцій.
Ініціація скорочення йонами кальцію розвивається так. Йони кальцію, що проникли усередину клітини, впливають на регуляторний білок кальмодулін (замінює тропонін-тропоміозинову систему посмугованих м'язів), який активує кінази легких ланцюгів міозину. Внаслідок цього фосфатна група разом із відповідною кількістю енергії з АТФ переноситься на міозин, який, з'єднавшись із актином, набуває АТФ-азної активності і зумовлює процес скорочення (роботу поперечних містків).
Поки Са2+міститься в клітині, вона перебуває у скороченому стані.
Коли вміст Са2+ у саркоплазматичпому матриксі знижується, актоміозиновий комплекс дисоціює і м'язова клітина розслаблюється.
Сила скорочення гладких м'язів хребетних практично така сама, як і скелетних м'язів (30-40 Н/см2), проте витрата енергії гладкими м'язами (за однакового напруження) у 100-500 разів менша, що, ймовірно, пов'язано з особливою організацією скоротливого акту гладких м'язів.
Тривалість скорочення. Поодиноке скорочення гладкої м'язової клітини триває кілька секунд. Висхідна фаза скорочення триває 1-2 с, а розслаблення — -10 с (у м'язах кишок), тобто порівняно з посмуговаиим м'язом скорочення гладкого м'яза відбувається у 20-50 разів повільніше. Це зумовлено низькою швидкістю взаємодії скоротливих білків у гладких м'язових клітинах через низьку активність їхньої АТФ-ази. У зв'язку з цим тетанічне скорочення гладких м'язів виникає при частоті стимуляції 0,5—3 за 1 с. проте не всі гладкі м'язи скорочуються повільно.
Тонус гладких м'язів. Гладкі м'язи, входять до складу стінки внутрішніх порожнистих органів (шлунка, кишок тощо), можуть перебувати у скороченому стані годинами, а гладкий м'яз матки савців взагалі скорочений майже все життя без будь-яких ознак втоми, за винятком відносно коротких періодів вагітності.
Очевидно, що таке тривале скорочення не може бути тетанічним. Воно є тонічним скорочєнням, природу якого ще до кінця не з'ясовано. Ймовірно, тонічне скорочення гладких м'язів підтримується сталою високою концентрацією Са2+ у саркоплазмі м'язових клітин, яка створюється завдяки постійно відкритим кальцієвим каналам.
Тонічне скорочення гладких м'язів може змінюватись: стінка порожнистого органа у стані деякого розслаблення або скорочення може перебувати протягом певного часу. Це явище називають пластичним тонусом, воно властиве багатьом гладким м'язам і відіграє важливу роль у стабілізації тиску всередині органа. Так, тиск у шлунку чи сечовому міхурі мало змінюється в міру наповнення чи випорожнення цих органів.