

МІНІСТЕРСТВО АГРАРНОЇ ПОЛІТИКИ ТА ПРОДОВОЛЬСТВА
ДНІПРОПЕТРОСЬКИЙ ДЕРЖАВНИЙ АГРАРНИЙ УНІВЕРСИТЕТ
Кафедра загального землеробства та ґрунтознавства
ТЕХНІЧНА МІКРОБІОЛОГІЯ
Методичний посібник для студентів
За напрямом підготовки
6.0 51701 «Харчова технологія та інженерія»

ЧАСТИНА I
ЗАГАЛЬНА МІКРОБІОЛОГІЯ
ВСТУП
Мікробіологія (греч. micros — малий, bios — життя, logos — вчення)— наука про дрібні, невидимі неозброєним оком організми. Вона вивчає морфологію,систематику, фізіологію і біохімію, генетику, екологію мікроорганізмів, їхню роль і значення в кругообігу речовин, досліджує загальні умови їхньої життєдіяльності і способи спрямування цієї діяльності на користь людині. Мікробіологія вивчає - бактерії, міксобактерії, мікоплазми, рикетсії, віруси, фаги, спірохети, актиноміцетів, цвілеві гриби і дріжджі. До мікроорганізмів відносяться також найпростіші та мікроскопічні водорості. Різноманіття мікроскопічних істот виправдовує існування у складі мікробіології особливих наук, які вивчають окремо різні групи мікроорганізмів. Так, бактерії вивчає бактеріологія, мікроскопічні гриби - мікологія, самостійною наукою стала вірусологія.
Задачі мікробіології в народному господарстві настільки різноманітні, що існує галузеве розподілення її на ряд спеціалізованих наукових дисциплін (сільськогосподарська мікробіологія, ґрунтова, медична, ветеринарна, геологічна, водна, технічна мікробіологія та ін,.).
Загальна мікробіологія вивчає основні закономірності розвитку та життєдіяльності всіх груп мікроорганізмів, а також роль та значення мікробів в процесах кругообігу речовин в природі з метою можливості регулювання останніх в інтересах людини.
Вивчення мікробного складу ґрунту – першочергова задача ґрунтової мікробіології, так як, не знаючи видового складу та кількості мікроорганізмів, не можна планомірно вирішувати багатьох проблем ґрунтової родючості.
Важливим є встановлення так званої ризосферної (кореневої) мікрофлори, що має великий вплив на живлення та ріст рослин. Мікроорганізми утворюють гумус ґрунту, оструктурюють його, без них немислиме первинне формування ґрунтів, немає родючості ґрунту.
Загальновідома роль бульбочкових бактерій, які, знаходячись в симбіозі з бобовими рослинами, поліпшують їх азотне живлення та збагачують грунт азотом.
Великим досягненням ґрунтової мікробіології є виявлення у мікроорганізмів біологічно активних речовин (антибіотики, вітаміни, гормони, амінокислоти, ферменти і т.д.), що грають особливу роль в житті вищих та нижчих організмів. Вчення про синтез цих речовин мікробами складає особливий розділ мікробіології.
Технічна мікробіологія розробляє наукові основи використання біохімічної діяльності мікроорганізмів у різних виробничих процесах , а саме - для розвитку хлібопечення, виробництва молочнокислих продуктів, пивоваріння, винокуріння, отримання ацетону, бутилового спирту, лимонної та оцтової кислоти та інших технічних продуктів. Технічна мікробіологія розробляє технологію виробництва органічних кислот, спирту, вина, пива, замочування прядильних культур, внесення бактеріальних добрив і засобів захисту рослин; серед її завдань виділяють також розроблення методів боротьби з корозією металів тощо.
Важливою задачею медичної та ветеринарної мікробіології є вивчення хвороботворних мікроорганізмів та їх взаємодії з організмом людини або тварини, а також розробка методів специфічної діагностики, профілактики та лікування інфекційних захворювань.
Серйозна увага приділяється розробці та використанню ефективних методів застосування мікроорганізмів в тваринництві з метою отримання кормових продуктів (амінокислот, білків), виробництва кормових антибіотиків, вітамінів, ферментів і т.д.
РОЗДІЛ 1. СИСТЕМАТИКА, МОРФОЛОГІЯ І УЛЬТРАСТРУКТУРА МІКРООРГАНІЗМІВ
Для більшості представників мікросвіту характерною є одноклітинна будова. Бактеріальна клітина оточена оболонкою ( а в деяких випадках – цитоплазматичною мембраною) і містить цитоплазму па інші органели. За основними типами клітинної будови всі живі організми поділяються на дві великі групи: прокаріоти і еукаріоти.
ПРОКАРІОТАМИ ( PROCARYOTAE) називають мікроорганізми, які не мають чітко диференційованого ядра, а містять його аналог – нуклеоїд. До них належать бактерії різних систематичних груп, синьозелені водорості, які називають ціанобактеріями. Вважають, що прокаріоти – похідні первісних організмів, які існували понад три мільярди років тому. До прокаріотів
відносяться і недавно відкриті архебактерії.
ЕУКАРІОТАМИ (EUCARYOTAE називають всі одноклітинні та багатоклітинні організми, які мають сформоване ядро, відмежоване від цитоплазми ядерною мембраною. До еукаріотів, що їх вивчають у мікробіології, в тому числі технічній мікробіології,відносяться гриби, водорості, найпростіші. За будовою вони подібні до рослинних та тваринних клітин
Дослідниками ультраструктури клітин з’ясовано глибокі фундаментальні відмінності у будові прокаріотичних і еукаріотичних клітин Прокаріотична клітина має тільки одну внутрішню порожнину, яка утворена клітинною мембраною. У прокаріотичній клітині відсутні оточені ліпопротеїдними мембранами компартменти або органели, тобто вона не має замкнутих вторинних порожнин. У клітинах прокаріотів ядерний апарат не відмежований ядерною оболонкою з поровим комплексом від цитоплазми.У них відсутні мітохондрії і хлоропласти, ендоплазматична сітка, диктіосоми, лізосоми і мікротільця, мікрофіламенти, тубулінові мікротрубочки і мітотичний апарат. Геном у цих мікроорганізмів являє собою кільцеву хромосому, яка складається з ДНК.
В еукаріотичних клітинах, на відміну від прокаріотних, є вторинні порожнини, ядро, оточене подвійною мембраною, утворює вторинну порожнину. В цитоплазмі еукаріотів є також інші двомембранні органели: мітохондрії, пластиди,до одно мембранних відносяться: апарат Гольджі, ендоплазматична сітка, лізосоми, вакуолі та ін..
Форма та розміри бактерій
Бактерії ( від. лат.bacteria- паличка) відзначаються простотою форми. За зовнішнім виглядом їх поділяють на такі основні групи: кулясті (коки), паличкоподібні ( бактерії, бацили, клострідії), звивисті ( вібріони, спірили, спірохети, нитчасті (сіркобактерії,залізобактерії тощо), бактерії незвичайних форм.
Коки(лат.coccus –зерно).
Кулясті бактерії –коки-бувають сферичні, еліпсоподібні, бобовинні, ланцетоподібні. За характером поділу, розміщенням й біологічними властивостями вони поділяються на мікрококи, диплококи, стрептококи, тетракоки, сарцини і стафілококи.
1. Мікрококи (Micrococcus)-коки, які після закінчення, поділу, не залишаються разом, а розташовуються безладно, по одному: 2. Диплококи (грец. diplos - подвійний)-коки, які після поділу клітини залишаються з'єднаними попарно. 3. Тетракоки (грец. tetra - чотири)-коки, зцеплені по чотири внаслідок поділу клітини послідовно в двох взаємно перпендикулярних напрямках. 4. Сарцини (від лат. Carcio - з'єдную)-скупчення коків у вигляді пакетів з 8-18 і більше клітин, коли вони діляться постійно в трьох взаємно перпендикулярних площинах.
5. Стрептококи (грец. streptos - ланцюг)-коки, у яких поділ відбувається завжди в одному напрямку, а виникаючі нові клітини не відокремлюються одна від одної і утворюють різної довжини ланцюги. 6. Стафілококи (грец. staphyle - гроно) - коки, розташовані у вигляді грон винограду, завдяки поділу клітин в різних напрямках.
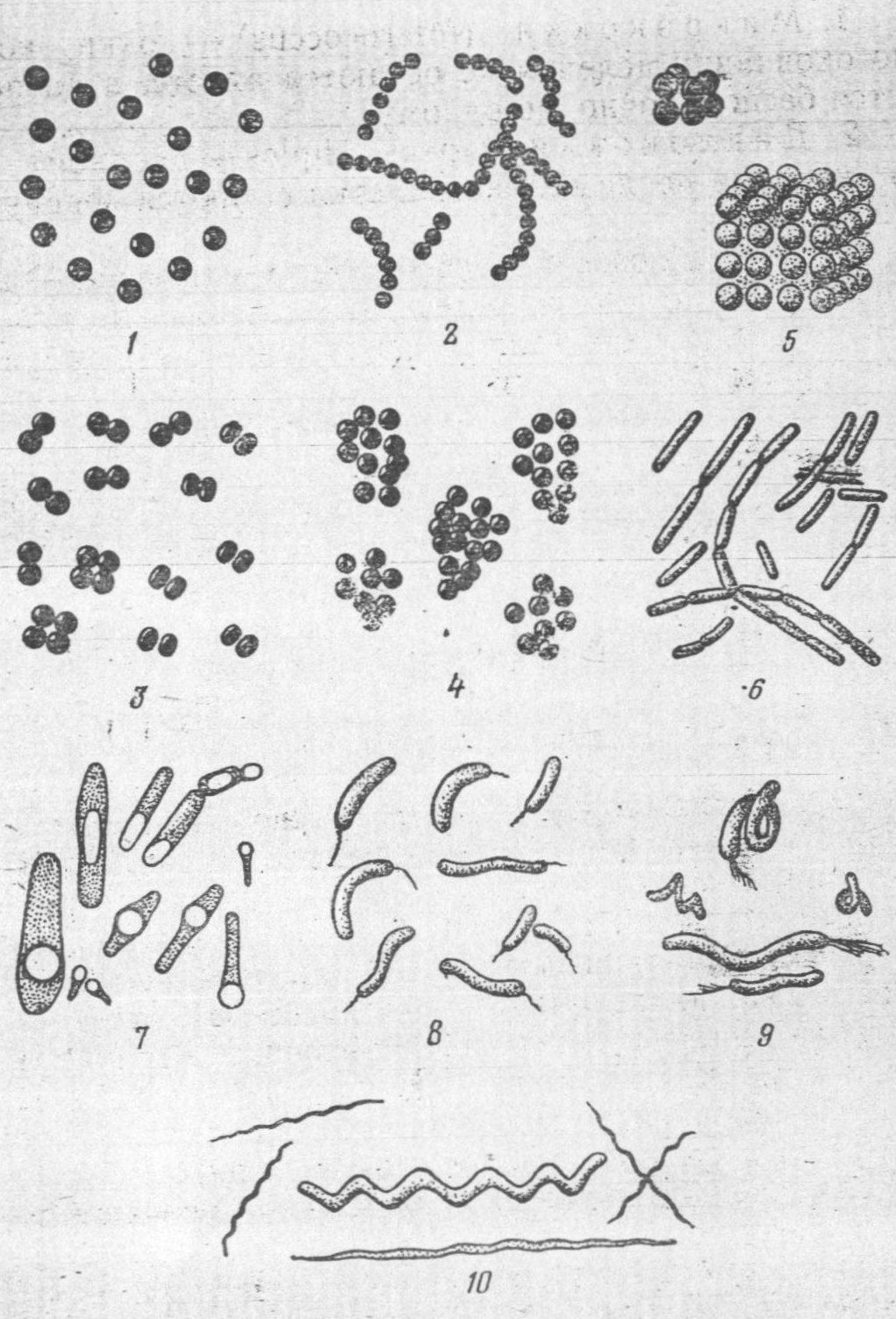
Палочковидні бактерії. Клітини їх можуть бути короткими і довгими. Кінці клітин бувають зрізаними, злегка втиснутими всередину, закругленими або загострені, іноді на них спостерігаються здуття. Ці бактерії прийнято ділити на дві групи: палички, утворюючі спори, що називаються бацилами (від лат. bаcillum - паличка), а палички, що не утворюють спор, - бактеріями (від грец. Bacteria - паличка).
Рис.3.
Схема будови бактеріальної клітини
(по В.І.Бірюзовій):
Н
– нуклеоїд; М – мезосоми; АМ – аналоги
мітохондрій; КС – кліткова стінка; К –
капсула; ЦМ – цитоплазматична мембрана;
АЦ – аналоги центросом; В – вакуоля;
ВП – видільні пухирці; Б – блефаробласти;
ЖГ – джгутики; Ж – жир; Вол. – волютин;
Г – глікоген; Р – рибосоми.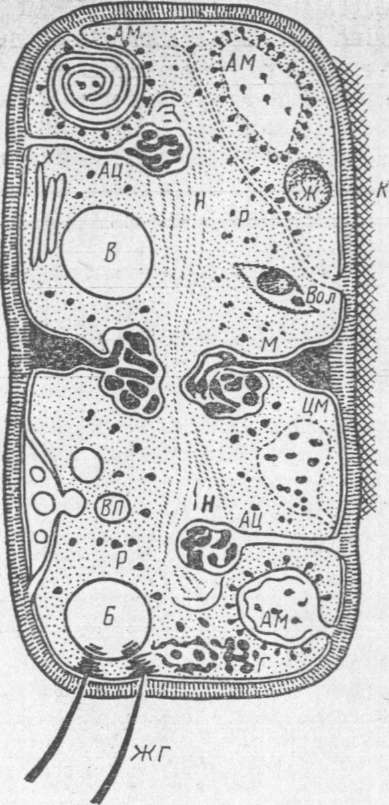
Це група одноклітинних, ниткоподібних, спірально звивистих мікроорганізмів розміром від 7 до 500 мкм в довжину і від 0,3 до 1,5 мкм в поперечнику. Тіло спірохет складається з осьової нитки, навколо якої спірально завита цитоплазма, і покрито тонким цитоплазматичним шаром - перипластом. Спірохети не мають оформленого ядра, цитоплазма у них гомогенна, клітинна стінка відсутня. Розмножуються поперечним поділом, спор не утворюють. У деяких видів на кінці тіла спірохети розташовується пучок джгутиків. Окремі види хвороботворні для людини і тварин (збудники сифілісу і лептоспірозу). За будовою ця група мікроорганізмів займає проміжне положення між бактеріями і найпростішими.
Розміри бактерій. Величина їх вимірюється в мікрометрах (мкм) і нанометрах (нм). Діаметр клітин у кокових форм варіює від 1,5 до 2,5 мкм. Звичайна величина паличкоподібних бактерій знаходиться в межах 0,8-1,2X2-10 мкм. Серед звивистих форм, а також у сіркобактерій і залізобактерій зустрічаються більш крупні клітини, розміри яких сягають до 2-3X20-50мкм .Проте серед мікроорганізмів є свої «гіганти», наприклад,спірохети можуть мати довжину 500 мкм завдовжки. У найменших бактерій –мікоплазм діаметр клітини становить 0,12 – 0,15 мкм.
Субмікроскопічна будова бактеріальної клітини.
За сучасними даними, бактеріальна клітина складається з тих же морфологічних компонентів, що і клітини тварин і рослин: вона має оболонку, цитоплазму, ядерний апарат і цитоплазматичні включення (рис. 3).
Оболонка клітини тонка, відносно щільної структури, товщиною від 10 до 60 нм, складає біля 20% маси сухої речовини.
Клітинну оболонку(стінку) можна виявити за допомогою спеціального забарвлення. В незабарвленому вигляді вона помітна тільки у великих бактеріальних форм, наприклад у залізобактерій і сіркобактерій. Повне уявлення про оболонку можна отримати за допомогою електронного мікроскопу.
Оболонка надає клітині певну форму, регулює проникнення в неї поживних елементів, виділення назовні продуктів обміну, ферментів та інших речовин. Оболонка захищає клітину від дії зовнішнього середовища і витримує значний внутріклітинний осмотичний тиск 300-600 кПа.
Залежно від особливостей хімічного складу оболонки всі види бактерій по-різному ставляться до способу забарвлення, розробленому датським вченим Грамом. За цим способом забарвлення бактерії можуть бути поділені на грампозитивні, що стійко забарвлюються у фіолетовий колір, і грамнегативні, що не зберігають цього забарвлення. Сутність забарвлення по Граму полягає в тому, що бактеріальну клітину обробляють генцианвіолетом та йодом, а потім промивають спиртом. Клітинна оболонка грампозитивних бактерій містить значну кількість мукополісахаридних, поліфосфатнуклеотидних комплексів, з якими генціанвіолет і йод міцно зв'язуються і тому повільно вимиваються спиртом. У грамнегативних бактерій зазначених вище комплексів значно менше, і вони розміщуються в глибині клітини. Тому генціанвіолет і йод легко руйнуються спиртом і клітинна оболонка швидко знебарвлюється.
Оболонка складається з трьох шарів: слизового шару, клітинної стінки і цитоплазматичної мембрани.
Рис.4.
Капсули у бактерій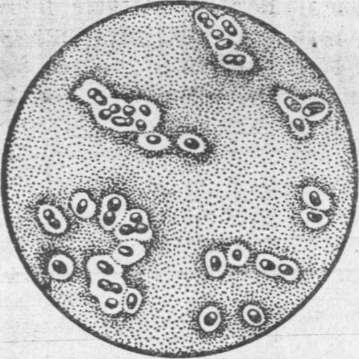
Капсульна речовина складається з полісахаридів і поліпептидів. В окремих видів бактерій капсули містять азот, a Acetobacter xylinum-єдиний вид, який має капсулу, що складається з целюлози.
Клітинна стінка - складається з полісахаридів та протеїдів. Під електронним мікроскопом на ультратонких зрізах бактерії стінка видна у вигляді одношарового утворення, більш щільного, ніж вміст бактеріальної клітини. .На клітинну стінку припадає від 5 – 50% сухої речовини клітини. До складу клітинної стінки прокаріотів входить сім груп хімічних речовин
Головним структурним компонентом оболонок більшості бактерій є пептидоглікан (муреїн). Тільки у мікоплазм, L -форм бактерій, архебактерій та деяких метаноутворюючих і галофільних бактерій він відсутній. Пептидоглікан – гетерополімер,який складається з лінійних молекул глікану. Полісахаридний кістяк муреїну побудований із залишків,які чергуються, -ацетилглюкозаміну і -ацетилмурвмової кислоти, з’єднаних між собою .У грам позитивних бактерій нині виявлено близько 100 різних хімічних типів мурену. -1,4 глікозидних зв’язків.
До сітки пептидоглікану клітинної оболонки грампозитивних прокаріотів входять також тейхоєві і тейхуронові кислоти, поліпептиди ліпополісахариди, ліпопротеіни та ін..
У складі клітинної оболонки грам позитивних бактерій також виявлено в невеликих кількостях білки,ліпіди і полісахариди. З’ясовано, що полісахариди і ліпіди можуть ковалентно з’єднуватися з макромолекулами оболонки. Стосовно білків, то припускають, що вони виконують захисну функцію.
У грамнегативних еубактерій будова клітинної оболонки є набагато складнішою, ніж у грампозитивних.У середині оболонки цих бактерій міститься пептидоглікановий шар. Ззовні від нього розташований ще один шар(зовнішня мембрана), який складається з фосфоліпідів, ліпополісахаридів і білків, а під ним – цитоплазматична (внутрішня) мембрана, до складу якої також входять фосфоліпіди, білки тощо.
Між зовнішньою і внутрішньою мембранами клітинної стінки грам негативних бактерій існує периплазматичний простір, в якому, крім муреїного шару,містяться специфічні білки, олігосахариди, неорганічні речовини і вода. Периплазматичні транспортні білки відіграють важливу роль у надходженні в клітину амінокислот,цукрі, фосфатів,тощо.
Серед прокаріотів виявлено види бактерії , клітинна оболонка яких за структурою і хімічним складом помітно відрізняється від грам позитивних та грам негативних типів. Вони належать до групи архебактерій, зокрема оболонки метан утворюючих архебактерій містять пептидоглікан особливого хімічного складу. В інших представників цієї групи клітинна стінка складається переважно з кислого гетеро полісахариду. Оболонка екстремальних галофітів, метан утворюючих та ацидотермофільних архебактерій складається тільки з білка. Ці бактерії не забарвлюються за Грамом.
В залежності від будови клітинної стінки прокаріотичні мікроорганізми розподіляють на слідуючі відділи ((за Д. Бергі,1984).
ВІДДІЛ 1.Graciliscutes (грацілікути). До цього відділу належать грам негативні прокаріоти різних форм. Вони бувають рухомими і нерухомими, ендоспор не утворюють, розмножуються шляхом поділу, а деякі брунькуванням. Серед них є аероби і анаероби, фототрофи, хемотрофи, сапрофіти і паразити .Широко поширенні в ґрунтах водоймах,а також в живих організмах.Відділ поділений на три класи: скотобактерії, два класи фото синтезуючих організмів.
ВІДДІЛ 2. Firmicutes (фірмі кутні грампозитивні прокаріоти). В цьому відділі об’єднано мікроорганізми, які мають кулясту, паличкоподібну, нитчасту, розгалужену та інші форми. Серед них є рухомі і нерухомі, аеробні,анаеробні, спороносні і не спороносні організми .Переважна більшість з них розмножується бінарним поділом, а деякі спорами. З огляду на морфологію, ці прокаріоти запропоновано ділити на два класи: фірмі- -бактерії і талобактерії, розгалужені форми.
ВІДДІЛ 3.Tenericutes (тенерикути). До цього відділу належать грам негативні прокаріоти. які не мають клітинної оболонки і не синтезують пептидоглікану. Клітини їх оточені цитоплазматичною мембраною, а тому за формою вони є дуже плеоморфними. Тенерікути розмножуються бінарним поділом,брунькуванням та фрагментацією. Серед них є сапрофіти, паразити, патогенні організми – збудники захворювань рослин, тварин,людини. Відділ включає клас – мікоплазми.
ВІДДІЛ 4.Mendosicutes (мендозикутні прокаріоти). У більшості мендозикутів в клітинних стінках немає пептидоглікану. Вона може бути побудована тільки з білка або гетеро полісахаридів. Серед них переважно облігатні анаероби, не утворюють ендоспор, забарвлюються за Грамом негативно і позитивно. Ці організми характеризуються різноманітністю метаболізму і здатністю жити в різних екстремальних умовах. В цьому відділі об’єднано прокаріоти різних форм, які за походженням вважаються старшими за представників 1 і 2 відділів. Відділ включає клас – архебактерій.
Цитоплазматична мембрана. До внутрішньої поверхні, клітинної стінки щільно прилягає зовнішній шар цитоплазми-цитоплазматична мембрана (пері-пласт), що має вид тонкої блискучої плівки. Мембрана складає понад 10% маси сухої клітини і складається з ліпопротеїнових і рибонуклеопротеїнових комплексів. Товщина її не перевищує 10 нм.
Мембрана служить місцем інтенсивної фізіологічної активності, так як є носієм багатьох ферментів.
Мембрана захищає цитоплазму. Крім того, вона підтримує постійний внутрішньоклітинний осмотичний тиск, затримує в цитоплазмі поживні речовини і солі і одночасно сприяє виділенню продуктів обміну. Цитоплазматична мембрана володіє виборчою проникністю, тут відбувається ензиматична обробка речовин для засвоєння їх - клітиною.
У різних груп прокаріотів цитоплазматична мембрана утворює локальні впинання, які називаються мезосомами. Ці мембранні утворення добре розвинуті у грампозитивних еубактерій.У грам- негативних вони трапляються рідше і менш розвинені. За формою, розмірами і розміщенням розрізняють три основні типи мезосом:ламелярні (пластинчаті), тубулярні (трубчаті), везикулярні (пухирчасті). Мезосоми найчастіше містяться в зоні формування клітинної перетинки і поділу бактеріального ядра. Можливі функції мезосом :відіграють певну роль в поділу клітини,виконують структурну та метаболічну .
Цитоплазма у бактерій являє прозору водянисту або злегка в'язку однорідну суміш колоїдів. В ній можуть бути суспендовані також пігменти, запасні поживні речовини і жири. За хімічним складом цитоплазма являє собою складну суміш води; білків, нуклеїнових кислот, вуглеводів, ліпідів, мінеральних речовин та інших органічних сполук. У молодих бактеріальних клітинах цитоплазма гомогенна, в клітинах старих культур з’являється зернистість.
В цитоплазмі бактерій відбуваються складні процеси обміну речовин, в результаті чого внутрішня структура клітини безперервно оновлюється.
З віком в цитоплазмі клітин з'являються різні включення. Більшість їх відноситься до резервних речовин:
Рибосоми – дрібні округлі рибонуклеопротеїнові гранули розміром 10 - 20 нм, розташовані довільно в цитоплазмі бактерій і складаються на 40% із білку і на 60% із РНК. ЇХ кількість в клітині залежить від інтенсивності синтезу білка і може коливатися від % 5000 до 90000. Рибосоми прокаріотів мають константу седиментації 70 ,від чого і дістали назву 70 S-частинок. Рибосоми є центрами синтезу білка в клітині. Підчас синтезу вони набувають форми агрегатів, які називаються полі рибосомами.
Ці полісоми містяться в цитоплазмі або зв’язані з мембранними структурами.
.
Рис. 5. Ультратонкі
зрізи клітин (під електронним мікроскопом)
1
— ядерна вакуоля;
2
— щільний тяж.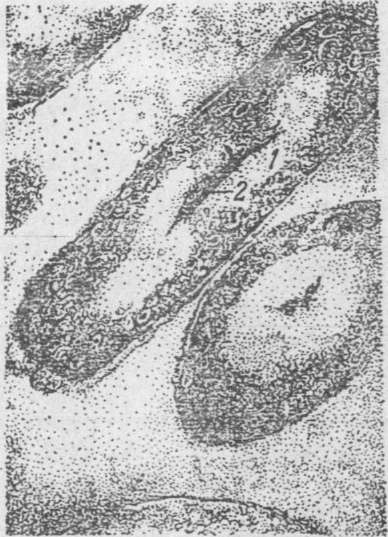
У істинних бактерій ДНК не оточена мембраною і зосереджена в обмежених ділянках у вигляді дискретних (переривчастих) формених структур, які найчастіше іменуються нуклеоїдами, або ядерним апаратом (рис. 5).
Електронна мікроскопія дозволила встановити, що область розташування ДНК бактеріальної клітини заповнена тонкими нитками і виглядає менш щільною, ніж навколишня цитоплазма. ДНК бактерій розглядається як хромосома і являє собою нитку, замкнуту в кільце. Нуклеоїд у прокаріот ще називають бактеріальною хромосомою.
Встановлено, що вміст вміст пар нуклеотидних основ у молекулі ДНК бактеріальної хромосоми А+Т і Г+Ц є постійним для даного виду організму. У молекулі ДНК нуклеоїда зосереджена майже вся генетична інформація прокаріотичної клітини.
У клітинах прокаріотів понад з бактеріальною хромосомою містяться і поза хромосомні кільцеві молекули ДНК, що дістали назву плазмідів. Вони здатні до автономної реплікації і зумовлюють деякі сподкові властивості бактерій, наприклад здатність до кон’югації , резистентність до антибіотиків.
Таким чином нуклеоїд- є носієм спадкових властивостей клітини і контролює синтез специфічних білків
Внутрішньоплазматичні включення. В цитоплазмі прокаріотів різних видів містяться також включення, що оточені білковою мембраною і функціонують як структури. До них належать хлоросоми, фікобілісоми, аеросоми, магнітосоми, карбоксисоми. Серед включень, у яких відсутня мембрана, трапляються ті, що є місцем запасання поживних речовин та концентрування продуктів клітинного обміну, які відкладаються всередині прокаріотичних клітин .За концентрацією вони бувають рідкі, тверді та газоподібні.
Хлоросоми –внутрішньоплазматичні включення, які беруть певну участь в процесі фотосинтезу зелених бактерій.Вони мають фору пухирців, завдовжки 100 – 150 нм і завширшки 25 – 70 нм, оточені одношаровою щільною білковою мембраною,завтовшки 2 – 3 нм. У хлоросомах міятяться бактеріохлорофіли. Хлоросоми розташовані біля цитоплазматичної мембрани.
Фікобілісоми-характерні для ціанобактерій. Як і хлоросоми, вони приймають участь у процесі фотосинтезу. Ці включення мають напівсферичну форму і розташовуються правильними рядами на зовнішній поверхні фотосинтетичної мембрани та містять водорозчинні пігменти білкової природи:фікобіліпротеїни.
Карбоксисоми-структури, що мають вигляд багатокутників, виявлені в клітинах фототрофних хемолітотрофних еубактерій. Вони оточені одношаровою мембраною білкової природи і містять фермент – рибулозодифосфаткарбоксилазу, який бере участь у процесі фіксації СО2 у відновному пентозофосфатному циклі.
До включень, які мають пристосувальне значення належать магнітосоми і аеросоми. Магнітосоми – виявлені в клітинах бактерій,для яких характерний магнітотаксис, тобто здатність плавати вздовж ліній магнітного поля. Вони представляють собою частинки комплексного оксиду двовалентного і тривалентного заліза, оточені білковою мембраною, можуть мати різну форму і місце розташування.
До включень, які виконують роль запасних поживних речовин,належать полісахариди, ліпіди, поліпептиди, полі фосфати тощо. Із включень полісахаридної природи в клітинах найчастіше відкладаються глікоген, крохмаль, гранульоза (крохмалеподібна речовина ). У випадку несприятливих умов ці сполуки використовуються прокаріотами як джерело вуглецю і енергії.
Ліпіди накопичуються в клітинах у вигляді гранул і крапельок жиру, зокрема таким включенням часто є полімер 0-оксимасляної кислоти. Як і полісахариди,ліпіди є для бактерій добрим джерелом вуглецю і енергії.
Дуже поширеними запасними речовинами у багатьох прокаріотів є полі фосфати, які дістали назву волютину (метахроматинові гранули).Волютин накопичується в оцтовокислих, молочнокислих, азот фіксуючих та інших видах бактерій. Гранули волютину складаються переважно із полі фосфатів, а тому його найчастіше розглядають як внутрішньоклітинний резерв фосфору. Він використовується клітиною як джерело фосфору і енергії.
Запасні поживні речовини поліпептидної природи,які містять аспарагінову кислоту і аргінін виявлені у ціанобактерій. Вони є резервом азоту в разі браку його в середовищі.
До включень, що їх відносять до продуктів клітинного метаболізму, належать виявлені в цитоплазмі деяких бактерій кристалоподібні включення білкової природи ромбоподібної, кубічної та інших форм. Ці утворення виявились дуже токсичними для гусениць шкідливих комах.
Для багатьох сіркобактерій характерне відкладання в клітинах молекулярної сірки як продукту клітинного обміну. Для аеробних тонових бактерій сірка є джерелом енергії, а для анаеробних фото синтезуючих сіркобактерій вона є донором електронів.
У деяких бактерій виявлено включення білкової природи, округлої форми, які заломлюють світло. Вони дістали назву -тілець. Формування їх визначається вірусними або плазмідними генами. Функції -тілець досі ще не вивчені.
Ріст і розмноження мікроорганізмів
Під поняттям ріст розуміють гармонійне збільшення всіх хімічних компонентів, з яких складається мікробна клітина. Ріст мікроорганізмів залежить насамперед від того, чи є в середовищі вода, поживні речовини, фізіологічно активні речовини тощо. Відомо, що ріст мікроорганізмів завершується їхнім розмноженням, яке виявляється у збільшенні кількості особин мікробної популяції на одиницю об’єму.
Всі бактерії, дріжджі, цвілеві гриби, найпростіші, рикетсії розмножуються шляхом ділення, коли з однієї клітини утворюються дві особини. У одноклітинних водоростей, найпростіших, цвілевих грибів і дріжджів встановлений як статевий, так і безстатевий спосіб розмноження. Бактерії ж розмножуються переважно безстатевим поперековим поділом на дві рівні клітини (ізоморфне ділення), хоча іноді у них спостерігається і брунькування. Спадковість у бактерій існує, але передача. Швідкість розмноження бактерій залежить від складу середовища, температури, вологості і інших умов. При сприятливих умовах існування багато бактерій діляться через кожні 20-30 хв. Підраховано, що при такій швидкості розмноження одна клітина за 5 діб могла б дати кількість особин, здатних «заповнити басейни всіх морів і океанів (В.Л.Омелянський). Однак розмноження бактерій в природі обмежується різними несприятливими факторами зовнішнього середовища (дія сонячного світла, недостатня вологість, антагонізм між різними видами, нагромадження власних продуктів обміну в середовищі і ін.)
При розмноженні бактерій на рідкому поживному середовищі спостерігають кілька послідовних фаз:
1.Фаза затримки, або лаг-фаза, коли бактерії не розмножуються, пристосовуються до живильного середовища. Чим сприятливіше середовище для вирощуваної культури, тим коротше лаг-фаза.
2.Фаза логарифмічного розмноження, коли мікроорганізми енергійно розмножуються і число клітин збільшується в геометричній прогресії.
3.Стаціонарна фаза - число нових клітин які утворилися відповідає приблизно числу відмерлих. Кількість життєздатних бактерій залишається постійною.
4.Фаза старіння, коли припиняється розмноження і мікробні клітини через відсутність поживних речовин та накопичення продуктів життєдіяльності починають відмирати.
Відомо, що подовженість фаз залежить від штамових ознак, складу поживного середовища на якому вирощувалися мікроорганізми, кількості посівного матеріалу, різних фізичних та хімічних факторів.
Рухливість та процеси спороутворення у бактерій. Значна кількість видів паличкоподібних форм бактерій володіє активною рухливістю. Всі види спірил і вібріонів можуть самостійно рухатися. Коки нерухомі, за винятком окремих видів.
Переміщення в рідині здійснюється за допомогою джгутиків - тонких, довгих, звивистих ниток складної білкової природи. Вони відходять від щільного тільця в цитоплазмі і проникають через оболонку клітини. Довжина джгутиків зазвичай 6-9 мкм, але у деяких видів досягає 30 мкм і більше, у багато разів перевищуючи довжину бактерії. Товщина джгутиків коливається від 0,02 до 0,04 мкм. Під оптичним мікроскопом вони видні тільки після спеціального фарбування. За допомогою електронного мікроскопу можна детально розглянути форму, локалізацію і будову джгутиків у різних видів. За характером розташування джгутиків та їх кількості прийнято під рухливі бактерії ділити на групи: монотрихи – бактерії з одним джгутиком, розташованим на полюсі клітини, наприклад холерний вібріон; лофотрихи – бактерії з пучком джгутиків на одному з кінців клітини, наприклад флуоресціююча паличка; перитрихи - бактерії з великим числом джгутиків, розташованих по всій поверхні бактеріальної клітини, наприклад кишкова паличка (рис. 6).
При тривалому вирощуванні на твердих живильних середовищах рухливі бактерії можуть втратити джгутики, а на рідких - знову знайти їх.
Бактерії, у яких джгутики знаходяться на одному кінці клітини, переміщаються по прямій лінії. У перитрихів рух супроводжується жвавим перекиданням клітини в різні боки.

Рис.6. Джгутики у бактерій
Не слід змішувати самостійний рух бактерій з їх пасивним переміщенням в результаті молекулярного руху зважених у воді найдрібніших частинок, названим броунівським. Броунівський рух характеризується безладним коливанням бактерій на одному місці. У більшості монотрихів і лофотрихів швидкість руху досягає 50-60 мкм в секунду, у перитрихів - 25-30 мкм.
Спороутворення у бактерій.
Спороутворення у бактерій є одним із засобів пристосування їх до несприятливих зовнішніх умов (нестачі поживних речовин, наявності шкідливих продуктів обміну в навколишньому середовищі, занадто високої чи занадто низької температури та ін.)
Спороутворення характерно для паличкоподібних форм, серед кокових форм воно зустрічається рідко (Planosarcina ureae), а у звивистих форм взагалі не спостерігається.
На відміну від вегетативних форм, коли в клітині енергійно виявляються всі фізіологічні процеси, спори - це форма бактерій, що покоються (анабіоз). Спори у різних видів бацил відрізняються один від одного за формою, розміром і розташуванням в клітині. Так, спори сінної палички мають овальну форму і розташовуються в центрі клітини, спори у правцевій клітини кругла, розташована на кінці, і діаметр її більше діаметра вегетативної клітини (форма барабанної палички). Іноді велика спора утворює як би здуття в середній частині вегетативної клітини, що надає їй форму веретена (клостридіум) (рис.7).
Процес спороутворення у бактерій триває від декількох годин до діб. Він починається з ущільнення вмісту клітини, в результаті чого утворюється проспора. До неї переходить весь вміст в клітині ДНК. Коло проспори цитоплазма ущільнюється та перетворюється в оболонку спори, що складається з двох шарів – зовнішнього та внутрішнього. Зовнішній шар більш щільний та погано проникний для води. Потім залишки вегетативної клітини руйнуються.
Рис.7.
Спори у бактерій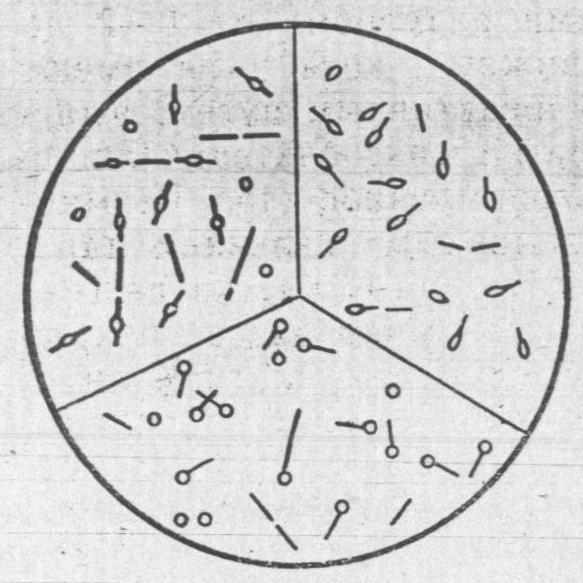
Бактерії у вигляді спор можуть зберігати життєдіяльність в стані спокою десятки і сотні років.
Потрапивши в сприятливі умови, спора починає проростати. Спочатку вона набухає за рахунок зростання клітини і поглинання води, збільшуючись майже в 2 рази, потім всередині спори посилюється активність ферментів, здатних гідролізувати оболонку спори, і в утворившийся отвір виходить назовні проросток, який потім перетворюється на нову вегетативну клітину. Проростання спори здійснюється протягом декількох годин (4-5). Поява проростка частіше спостерігається на одному з полюсів спори.
Морфологічна диференціація інших мікроорганізмів
Міксобактерії.Особливу групу складають міксобактерії, або слизисті бактерії (грец. muxo - слиз). Більшість міксобактерій - сапрофіти. Клітини їх мають вигляд паличок з загостреними кінцями розміром-від 1-2 до 10 мкм. Міксобактеріі рухливі, хоча і не мають джгутиків, вони пересуваються реактивним шляхом, виокремлюючи слиз в одну сторону. Клітини міксобактерій позбавлені щільної оболонки, мають оформлене ядро. Розмножуються перетяжкою з одночасним поділом ядра. В більш старих культурах вегетативні клітини стискаються і перетворюються в округлі мікроцисти, вкриті слизом, які, підносячись над щільним живильним середовищем, утворюють плодові тіла. При потраплянні в сприятливе середовище мікроцисти проростають Види міксобактерій відрізняються один від одного кольором, формою плодових тіл і розміром клітин.
Міксобактеріі - облігатні аероби, живуть і розмножються тільки при доступі вільного кисню. Вони воліють до нейтрального середовища, хоча переносять кисле і лужне з високою вологістю. Оптимальна температура для їх розвитку 30°С. Зустрічаються в ґрунті, гної, на рослинних рештках що розкладаються у вигляді слизових утворень. Вони активно руйнують найрізноманітніші речовини тваринного і рослинного походження, в тому числі клітковину, хітин, перетворюючи їх в з’єднання, придатні для харчування рослин.
Мікоплазми та L – форми .На відміну від бактерій мікоплазми і L-форми не мають клітинної стінки, внаслідок чого у них спостерігається виражений поліморфізм. Вони можуть бути представлені шароподібними, гіллястими тілами або, безформною зернистою масою. Розміри їх від 1-8 мкм-до форм що фільтруються (250 нм).
Мікоплазми вперше були відкриті в 1898р.. як найдрібніші мікроорганізми, здатні проходити крізь бактеріальні фільтри.
L-форми відкриті в 1935р. як варіанти мікроорганізмів, які втратили клітинну оболонку, але зберегли здатність росту і розмноження. L-форми описані майже у всіх хвороботворних бактерій.
За хімічним складом мікоплазми і L-форми відрізняються від бактерій відсутністю компонентів, що входять до складу клітинної оболонки.
Характерною загальною ознакою мікоплазм і L-форм є наявність цитоплазматичної мембрани товщиною 7,5 нм, за своєю будовою близькою до мембрани бактерій, а також ядерного і рибосомального апарату. Обидва види мікроорганізмів подібні за амінокислотним складом і за вмістом нуклеїнових кислот (РНК і ДНК) і вуглеводів. Мікоплазми і L-форми також схожі за формою і структурі колоній.
Дані про морфологію та біологію мікоплазм і L-форм дають підставу припускати, що мікоплазми є стабільними L-формами якихось бактерій, позбавлених клітинної оболонки і виникли в результаті генетичних мутацій, а стабільні L-форми бактерій є однією із стадій філогенезу-мікоплазм (В. Д. Тімаков).
Рикетсії.Рикетсії-своєрідні мікроорганізми, що займають проміжне місце між бактеріями і вірусами.
Рис.8.
Моделі вірусів:
1
– табакової мозаїки; 2 – поліомієліту.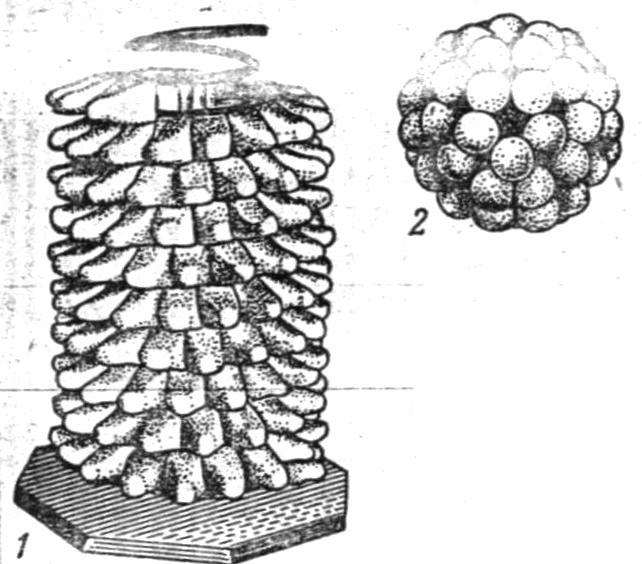
Рикетсії є збудниками ряду інфекційних захворювань у людини і тварин, типовим представником рикетсій є збудник висипного тифу.
Віруси.Віруси (від лат. Virus - отрута тваринного походження) - особливий клас дрібних мікроорганізмів, видимих тільки за допомогою електронного мікроскопа, що відрізняються від інших мікроорганізмів рядом морфологічних і біохімічних властивостей.
Були відкриті російським вченим Д. І. Івановським у 1892 р. При вивченні причин загибелі тютюну від мозаїчної хвороби. Івановський встановив, що заразний початок міститься в рослинному соку і проходить крізь бактеріальні фільтри (звідси назва «фільтрівні віруси»).
Віруси не мають клітинної структури. Розміри їх знаходяться в межах від 10 до 350 нм. Наприклад, розмір віруса ящеру 10 нм, а вірусу осповакцини – 200 – 350 нм.
За формою віруси можуть бути кулясті або еліпсоподібні, паличкоподібні, кубічні, ниткоподібні і булавовидні.
Найбільш проста вірусна частка (віріон) складається з нуклеопротеїду, в якому нуклеїнова кислота (РНК або ДНК) укладена в білкову капсулу - капсид. Капсид побудований з безлічі ідентичних білкових субодиниць, названих капсомірами (рис. 8). У більш складних вірусів нуклеокапсид має і оболонку, в якій містяться ензими (ферменти), що полегшують проникнення вірусу в сприйнятливу клітину. Деякі віруси містять також вуглеводи, ліпіди: і різні метали.
Віруси є внутрішньоклітинними паразитами, тобто здатні жити і розмножуватися тільки в клітині живого сприйнятливого організму. Всередину рослинних клітин віруси проникають крізь пошкодження, а в клітини тварини і людини - шляхом фагоцитозу або піноцитозу. Розмножуючись в клітках господаря, вони викликають їх загибель. При деяких вірусних інфекціях в уражених клітинах тканин виявляються особливі включення, характерні для певного вірусу. Так, при сказі в цитоплазмі нервових клітин головного мозку виявляються - тільця Бабеша - Негрі, при віспі в цитоплазмі клітин епітелію - тільця Гварнієри і т.д. Природа внутрішньоклітинних включень точно не з'ясована, вважають, що вони являють собою скупчення вірусних частинок або відмерлі частини кліток.
Деякі віруси отримані у вигляді кристалів (вірус тютюнової мозаїки). Кристалічний вірус складається з білка і нуклеїнової кислоти.
На фізичні та хімічні подразнення віруси реагують по-різному. Низькі температури не викликають загибелі вірусів. Плюсова температура 50-60°С впливає згубно. Віруси легко переносять висушування. Багато хто з них довго стійкі до дії етилового та метилового спиртів, до ефіру, а також до інших сильно впливаючих на бактерії хімічних речовин.
Фаги. Фаги (грец. phagein - пожирати)-віруси, що вражають бактеріальні клітини. У 1917 р.в Пастеровському інституті Д.Ерелль встановив явище лізису бактерій. Агент, що розчиняє бактерії, був їм названий бактеріофагом - «пожирачем бактерій», а процес лізису (розчинення) бактеріальних клітин - феноменом бактеріофагії. До Д. Ерелля феномен бактеріофагіі спостерігали і інші дослідники (Н.Ф.Гамалея, Творт). Крім бактерій, фаги виявлені і у актиноміцетів. Скрізь, де розмножуються бактерії і актиноміцети, виявляють і паразитуючих в них фагів.
Більшість фагів має своєрідну форму і складається з круглої голівки і довгого відростка (рис. 9). Розміри фагів колеблються: головки від 60 до 100 нм, відростка від 100 до 200 нм. Головка складається з білкової оболонки і укладеної в ній ДНК. Відросток фага має порожнистий стрижень, покритий зовні чохлом, що має здатність скорочуватися. На кінці відростку базальна пластинка з зубцями і нитками, які служать для адсорбції фага на бактеріальній клітині. Подібно до інших вірусів, фаги нерухомі. У результаті випадкових зіткнень їх з клітинами аги прикріплюються (адсорбуються) до поверхні останніх і крізь відросток вводять у клітину свою ДНК.
1
– ДНК+білок; 2 – зовнішня оболонка; 3 –
внутрішня оболонка; 4 – хвостовий шип.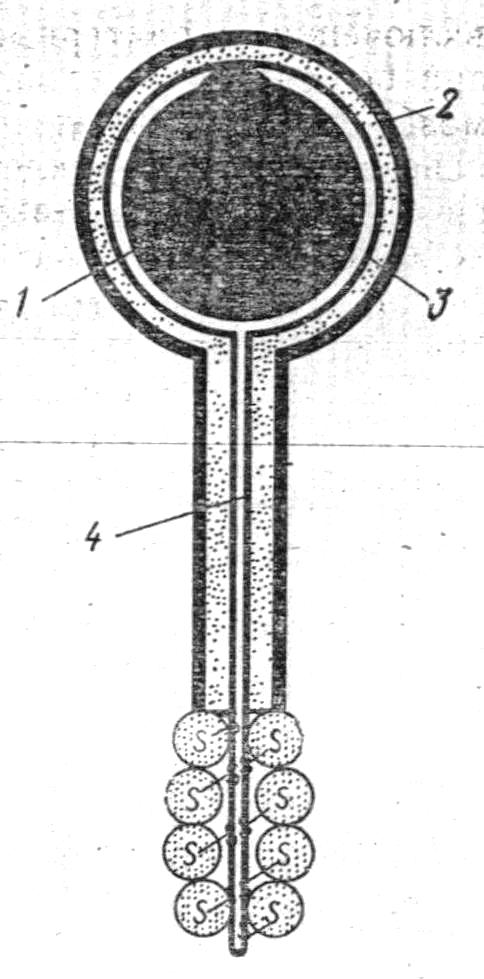 Рис.9.
Схема будови ага Т2:
Рис.9.
Схема будови ага Т2:
1. Адсорбція (фіксація)фага поверхнею чутливої мікробної клітини.
2. Проникнення вмісту фага (ДНК або РНК) в клітину.
3. Латентний період, коли в клітині йде інтенсивне утворення нових фагових частинок , кількість яких могла б досягати сотні або тисячі одиниць.
4. Лізис клітинної стінки бактерії під дією фагового лізоциму і потім вихід фагових частинок назовні.
Крім вірулентних фагів, що викликають лізис клітин, бактерії можуть заражатися недостатньо активними фагами, які не розмножуються в них і не викликають лізису. Такі фаги називаються помірними (профаги). А бактерії, що не лізуються від таких фагів, називаються лізогенними. При розмноженні бактерій ці фаги можуть переходити в дочірні клітини. Однак при певних умовах лізогенні культури бактерій можуть бути лізіровані фагами що в них знаходяться.
Фаги широко поширені в природі. Деякі використовуються в медицині та ветеринарії для профілактики інфекційних захворювань (дизентерія, паратиф, холера та ін.).
Найпростіші (Protozoa ). До найпростіших тварин відносяться мікроскопічні одноклітинні тварини, більш високо організовані в порівнянні з бактеріями. Вони досить різноманітні по величині, формі і структурі. У одних видів найпростіших оболонка добре відокремлена, міцна, грає захисну роль, в інших вона гнучка, тонка, є напівпроникною мембраною, а у деяких видів оболонка відсутня, і їх називають голими (амеба). Всі найпростіші мають здатність пересуватися за допомогою джгутиків, війок і псевдоподій.
Цитоплазма найпростіших являє собою колоїдну масу, що складається з двох шарів: зовнішнього, більш щільного, званого ектоплазма, і внутрішнього -ендоплазма. В цитоплазмі виявляються вакуолі і різні включення (волютин, глікоген, крохмаль та ін.)
У більшості найпростіших є одне ясно відокремлене від цитоплазми ядро, проте зустрічаються особи, які мають два або декілька ядер кулястої, овальної або неправильної форми. Розмножуються найпростіші поділом, а також статевим шляхом. Ці мікроскопічні істоти в грунті представлені трьома основними групами:
1.Корененіжки (Rhizopoda). Типовий представник-амеба, пересувається за допомогою псевдоподій. Розміри її 10-40 мкм.
2.Джгутикові (Flagelata). Рух у них здійснюється за допомогою одного або декількох джгутиків. Розміри 5-20 мкм.
3. Війчасті (Ciliata). Пересуваються за допомогою війок. Харчування у них відбувається крізь ротовий отвір. Мається й вихідний отвір. Розміри 20-80 мкм.При сприятливих умовах найпростіші прискорюють мінералізацію органічних залишків у грунті.
Актиноміцети (Actіnomycetes). Актиноміцети, або променисті гриби (грец. Myke - гриб, actis - промінь), займають проміжне положення між бактеріями і пліснявими грибами. Це одноклітинні мікроорганізми, що представляють собою тонкі розгалужені нитки - гіфи, 0,5-1,2 мкм в поперечнику і різної довжини. Більшість їх забарвлюється за Грамом позитивно (рис.10). Гіфи мають оболонку, цитоплазму із значною кількістю включень і ядерний апарат.
Деякі види актиноміцетів не утворюють гіф, клітини їх можуть бути паличкоподібної форми, найчастіше викривлені з невеликими бічними виростами, що нагадують гілки (рід Micobacterium).
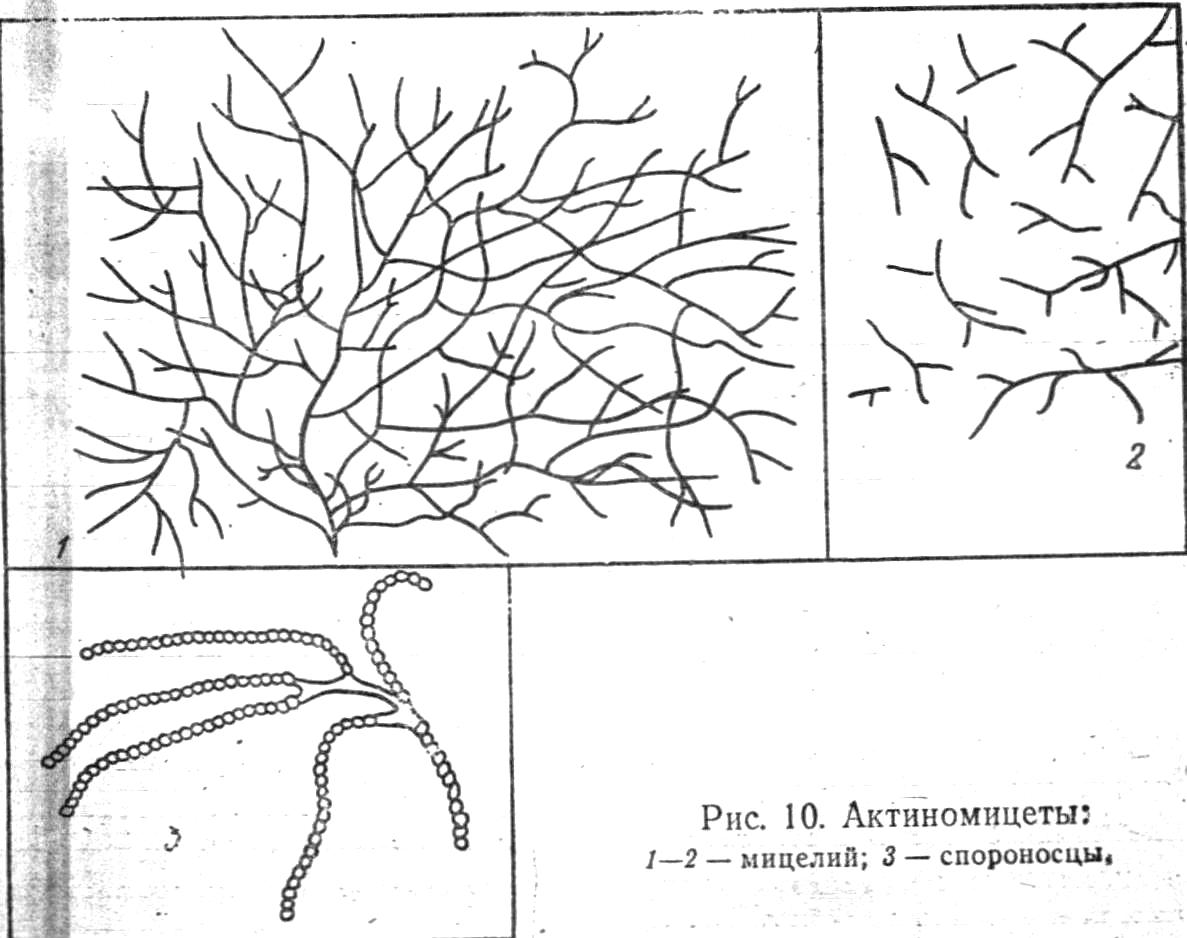
Розмноження відбувається шляхом розпаду міцелію на фрагменти-оідії, які за сприятливих умов можуть дати початок новому міцелію, а також спорами. Спори формуються на гілках повітряного міцелію (спороносцях). Вони легко відокремлюються від міцелію і, потрапляючи на поживний субстрат, швидко проростають. Окремі види актиноміцетів використовують для приготування антибіотиків (стрептоміцин, хлортетрациклін та ін.)
Таким чином, за будовою клітини, по відношенню до фарб і поживних середовищ актиноміцети подібні з бактеріями, а по характеру розмноження і проростання - з нижчими грибами.
Актиноміцети широко поширені в різних ґрунтах, у воді, гної та інших субстратах. Їм принад лежить велика роль у ґрунтоутворенні: вони посилюють розкладання органічних речовин в ґрунті і беруть участь у створенні гумусу. Багато видів актиноміцетів утворюють пігменти - червоний, чорний, бурий і ін.., роль яких поки невідома.
Ґрунтові актиноміцети переважно аероби (що розвиваються при доступі кисню повітря), але бувають і факультативні анаероби (життєдіяльність проявляється при відсутності вільного кисню). Оптимальна температура для розвитку актиноміцетів 23-37°С. Вони легко переносять висушування, можуть зберігатися в сухому ґрунті більше десяти років. Найбільш підходять для них ґрунти з рН 6,8-8,0. Чисельність їх у ґрунтах по відношенню до загальної мікрофлорі залежить від сезону року. Так, навесні вони складають 20%, восени – 30% і взимку -13%. У міру поглиблення в грунт кількість актиноміцетів зменшується.
Гриби(Fungi). Гриби - велика і різноманітна група рослинних гетеротрофних організмів, позбавлених хлорофілу. Гетеротрофний тип харчування обумовлює їх активну участь в розкладанні рослинних і тваринних залишків, завдяки чому вони грають важливу роль в утворенні органічних і гумусних речовин у ґрунті. Широке поширення грибів в ґрунті і значна активність пояснюються великою стійкістю їх до факторів зовнішнього середовища. Багато грибів добре розвиваються в умовах високої кислотності (рН 4 - 3), при якій затримується або повністю припиняється активна життєдіяльність більшості бактерій.
Гриби, як правило, аероби, тому розвиваються зазвичай на поверхні субстрату. Вони стійкі до висихання, але для розвитку воліють вологе середовище. Тіло більшості грибів складається з тонких ниток - гіфів, що сплітаються в міцелій (грибницю). Гриби, у яких гіфи розділені перегородками на клітини (септовані), називаються багатоклітинними, якщо ж гіфи не мають перегородок (не септовані), - одноклітинними. Будова клітин грибів мало чим відрізняється від будови інших мікроорганізмів. Клітини складаються з клітинної оболонки, цитоплазми і одного, двох або не декількох ядер. Цитоплазма містить вакуолі і різноманітні включення.
Деякі види грибів розмножуються вегетативно, коли відрізок міцелію з одного або великою кількістю клітин, потрапляючи на поживний субстрат, розростається і дає початок нової грибниці або утворює оідіі, після чого гіфи міцелію розпадаються на окремі клітини. Таку ж функцію виконує і міцелій,який брунькується у дріжджових грибів. Але найбільш характерним способом розмноження у грибів є утворення особливих спор. На певній стадії розвитку на поверхні міцелію з'являються особливі гіфи, що несуть спори (рис. 11). Зріла спора, потрапляючи на вологий субстрат що їй підходить, проростає і, поступово подовжуючись і розгалужуючись, утворює міцелій. У одних грибів спори виникають зовні (екзоспори), на спеціалізованих гіфах - конідієносцях. Такі спори називаються конідіями. Дозрілі конідії обсипаються. У інших грібів спори утворюються в кулястому спорангії, що сидить на кінці гіфи. Створювані в них спори (ендоспори) називаються спорангієспорами, а гіфи, що несуть спорангії, - спорангієносцями. У деяких грибів в спорангіях утворюються рухливі спори, забезпечені джгутиками, - зооспори. Зрілі спорангіі лопаються, і спори з них висипаються.
Гриби можуть розмножуватися і статевим шляхом. У цьому випадку після злиття двох ззовні однакових клітин з'являється нова клітина, яка розростається, покриваючись товстою багатошаровою оболонкою, і перетворюється в спору, яка називається зигоспорою. При злитті двох морфологічно різних (чоловічої та жіночої) клітин утворюється спора, що називається ооспора.
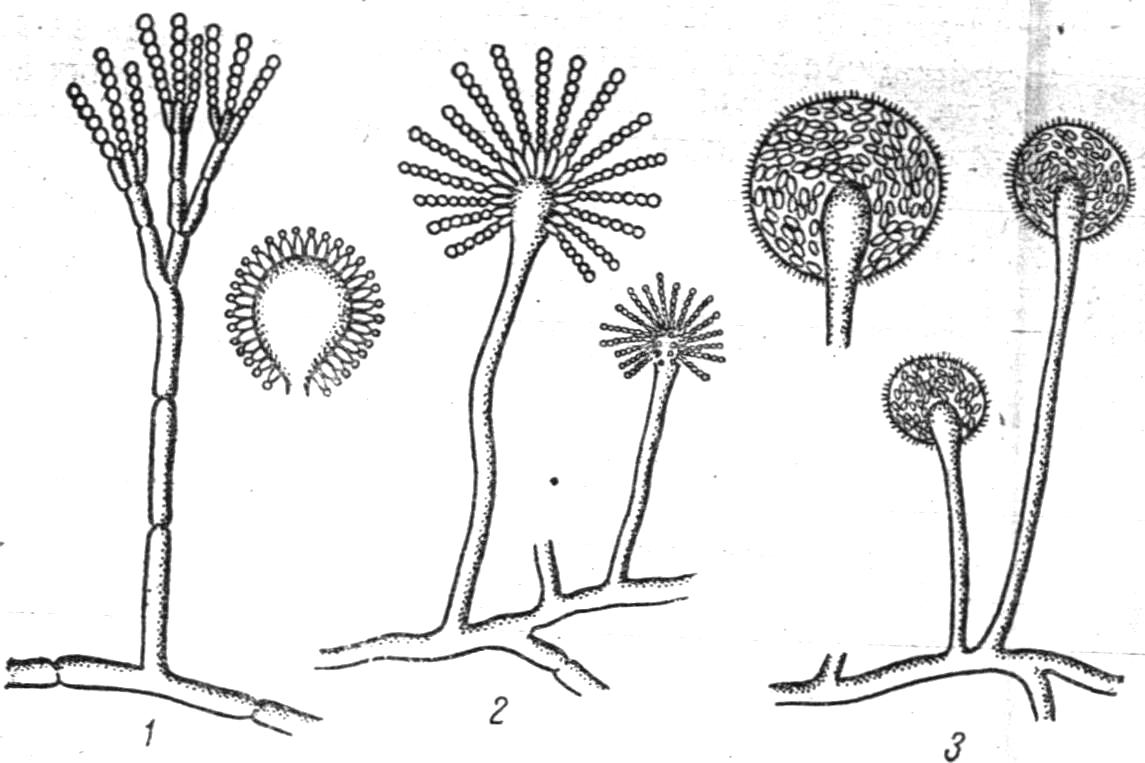 Рис.
11. Форми плодоношення у різних цвілевих
грибів! 1
- Aspergillus, 2 - Penicillium, 3 – Mucor.
Рис.
11. Форми плодоношення у різних цвілевих
грибів! 1
- Aspergillus, 2 - Penicillium, 3 – Mucor.
На підставі філогенетичних відносин всередині групи грибів, способів розмноження, морфологічних і фізіологічних ознак гриби розбиваються на класи, підкласи, сімейства, роди і види. За характером розмноження гриби поділяються на кілька основних класів. В основі класифікації лежать особливості їх будови та способи розмноження. Гриби розподіляються на нижчі та вищі. До нижчих відносяться: хітрідіоміцети (Chytridiomycetes), ооміцети (Oomycetes), зигоміцети (Zigomycetes). До вищих грибів відносяться: аскоміцети (Ascomycetes), базідіоміцети (Basіdiomycetes), недосконалі гриби (Fungi imperfecti).
Клас Хітрідіоміцети (Chytridiomycetes). У одних видів грибів цього класу є слаборозвинений міцелій, в інших тіло складається тільки, з голої грудочки цитоплазми. Розмноження переважно безстатеве - зооспорами. Більшість видів цього класу - внутріклітинні паразити нижчих і вищих рослин. Деякі з них вражають капустяну розсаду (чорна ніжка), бульби картоплі (рак).
Клас Ооміцети (Oomycetes). Ооміцети мають добре розвинений міцелій з багатоядерних несептированих гіф.
При статевому розмноженні в них утворюються ооспори. З патогенних грибів цього класу найбільш небезпечними паразитами є фітофтора і плазмопара.
Фітофтора, або картопляний гриб, вражає клубні та бадилля картоплі, а нерідко помідори і баклажани.
Плазмопара - гриб, який викликає хворобу віно граду, яка носить назву мілдью. Він вражає листя і ягоди.
Клас Зигоміцети (Zigomycetes). Зигоміцети утворюють одноклітинний міцелій. Типові представники- гриби родини мукорових (Мuсогасеае). Вони широко поширені в природі, зустрічаються з ґрунті, розвиваються на різних рослинних матеріалах і харчових продуктах у вигляді пухнастого сірого або темного нальоту. Мукорові гриби мають не септований міцелій, плодоносні гіфи з кулястим спорангієм нагорі, наповненим спорами, які утворюються в середині голівки (ендоспори).
При статевому розмноженні утворюються зигоспора. Безстатеве розмноження здійснюється нерухомими спорангієспорами.
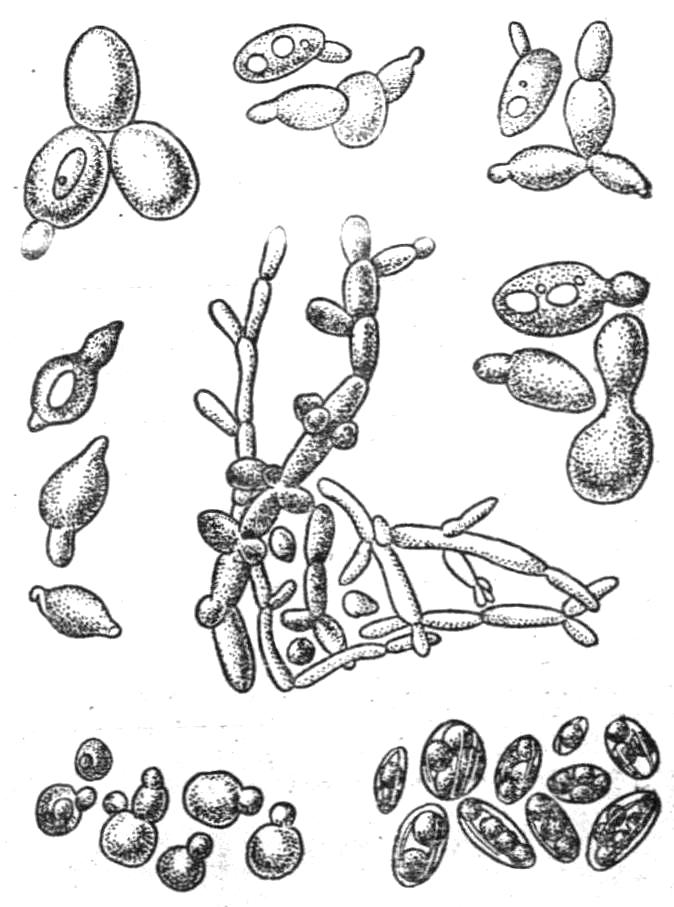 Рис.12.
Дріжджі
Рис.12.
Дріжджі
Дріжджі (Saccharomycetes). Одноклітинні неміцеліальні гриби. Клітини їх великі, кулястої або овальної форми, іноді циліндрові, лимоновидні (рис. 12), розміром 10 - 15 мкм, нерухомі. Дріжджова клітина складається з двухконтурної оболонки, цитоплазми і ясно вираженого ядра. В цитоплазмі виявляються 1-2 вакуолі, мітохондрії, а також включення - жир, волютин і глікоген.
Дріжджі розмножуються брунькуванням, рідше спорами і лише окремі види - простим поділом клітини. У деяких видів дріжджів встановлено статевий процес. Після з'єднання двох клітин, через канал відбувається злиття цитоплазми і ядер, в результаті утворюється одна велика клітина, в якій потім утворюється 18-20 спор. За способами розмноження і фізіологічними властивостям дріжджі ділять на порядки, сімейства, роди, види і раси. Дріжджові гриби розрізняють справжні та ложні. Останні відрізняються від справжніх тим, що не здатні утворювати спори.
Представники родини Saccharomycetaceae широко використовуються в спиртовій та пивоварній промисловості, а також при виготовленні вина і хліба (Saccharomyces сerevisiae, Saccharomyces ellipsoides). Поряд з корисними в цій групі зустрічаються види, що викликають псування сировини і харчових продуктів (Saccharomyces pasteurianum та ін.)
Вид дріжджів Torulopsis utilis застосовують для отримання харчових і кормових дріжджів. Torulopsis kefir використовують для виготовлення кефіру і кумису. Дріжджі роду Candida і деякі інші є збудниками мікозів тварин і людини.
Клас Базідіоміцети (Basіdiomycetes). Гриби з почленованим міцелієм. Основний спосіб розмноження - базидії з базидіоспорами (зазвичай 4). Більшість грибів цього класу живе на перегнійному ґрунті й рослинних залишках. Окремі види є паразитами і вражають польові, городні та садові рослини. З цієї групи найбільший інтерес для сільського господарства представляють будинковий гриб (Merulius), гриби, здатні давати на коренях рослин мікориз, - Boletus, а також сажкові та іржасті гриби.
Клас Недосконалі гриби (Fungi imperfecti). До цього класу належать багатоклітинні гриби, у яких не виявлено статевого процесу розмноження. Більшість грибів цього класу розмножується за допомогою конідій. Недосконалі гриби широко розповсюджені в природі. Деякі паразитують на культурних рослинах. Для агронома надають практичний інтерес такі роди, як фузаріум (Fusarium) і ботритіс (Botritis), викликають псування овочів, плодів, картоплі.
Водорості(Algae). Це велика група мікроскопічних рослин, що містять хлорофіл. Вони розповсюджені у воді, у вологих ґрунтах і беруть
активну участь в грунтовоутворюючих процесах. Водорості характеризуються мікроскопічними розмірами, складаються з однієї, декількох або багатьох клітин. Деякі водорості мають джгутики і активно пересуваються, подібно найпростішим.
Клітина водоростей вкрита оболонкою, є цитоплазма, одне або багато ядер (у синьо-зелених у водоростей диференційованого ядра немає), вакуолі і хроматофори - органи фотосинтезу, у водоростей відмічені три способи розмноження: вегетативне, безплідне і статеве.
Всі водорості в залежності від кольору пігменту можна підрозділити на три групи: зелені, синьо-зелені і діатомові.
Зелені водорості (Chlorophyceae ) пофарбовані в яскраво-зелений колір завдяки переважанню всередині клітин хлорофілу. З них в ґрунті найбільш часто зустрічаються наступні види: хламідомонада (Chlamydomonas) - одноклітинний організм з двома джгутиками, розташованими на передньому кінці клітини, особливо рясно розвивається навесні на вологому глинистому ґрунті; хлорококові (Chlorococcum)- ізольовані або утворюючі скупчення клітини, оточені слизовою муфтою, розмір 3-50 мкм.
Синьо-зелені водорості (Cyanophyceae) в деяких відносинах нагадують бактерії, але значно крупніші за них. Зазвичай в синьо-зелених водоростях містяться три пігменти: зелений пігмент - хлорофіл, блакитний - фікоціанін і червоний пігмент - фікоеритрин. В якості представників цієї групи можна назвати носток (Nostoc), осциляторію (Oscillatoria), хроококум (Chroococcum) (Рис. 13).
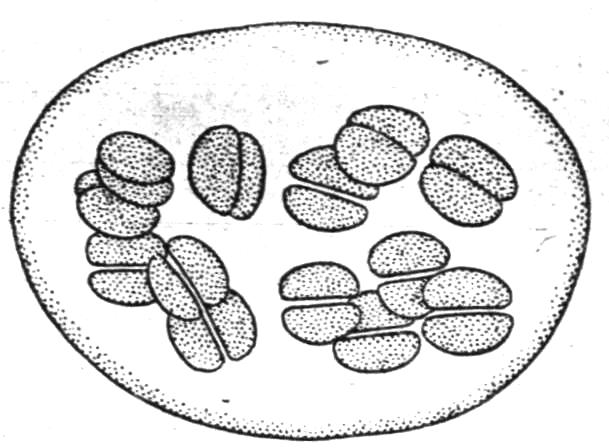 Рис.13.
Синьо – зелена водорость - Chroococcum
Рис.13.
Синьо – зелена водорость - Chroococcum
Водорості перших двох груп більш численні, і в сприятливих умовах кількість їх клітин може досягати 100 тис. в 1 г ґрунту. Встановлено, що, розвиваючись в ґрунті, водорості збільшують вміст у ній органічної речовини.
Відомо, що деякі види синьо-зелених водоростей здатні фіксувати атмосферний азот і перетворювати його в органічні сполуки, що вхдо складу їх цитоплазми. Особливо важливу роль водорості грають в підтримці родючості ґрунту рисових полів. Тут вони поповнюють запас кисню, знижують інтенсивність шкідливих відновних процесів, а окремі види фіксують за вегетаційний період до 15-20 кг азоту на 1 га.
Систематика і класифікація мікроорганізмів
Довгий час біологи вважали, що мікроорганізми не можна систематизувати, що серед них існує «хаос» (Лінней). Класифікації перших мікробіологів-систематиків О. Мюллера (1786), К. Еренберга (1838), Ф. Кона (1872), Мігула (1900) будувалися на морфологічних ознаках.
У сучасних системах класифікації враховують всі відомі ознаки мікроорганізмів: форму, розміри, забарвлення, рухливість і інші фізіологічні, культуральні та біохімічні властивості.
Для більшості сучасних визначників мікроорганізмів характерна відсутність єдиного принципу угрупування мікробів. Часто організми з одними і тими ж морфологічними ознаками відносяться до різних родів, сімейств і, навпаки, з неоднаковими морфологічними ознаками нерідко об'єднуються в одну групу.
Пояснюється це своєрідністю самих мікробів, примітивністю зовнішньої будови та розвитку, а також швидкою мінливістю під впливом умов зовнішнього середовища. Навіть такі морфологічні ознаки, як спороутворення, наявність, кількість і розташування джгутиків, структура клітини, бувають непостійними.
Не меншою мінливістю відрізняються і фізіологічні ознаки. Так, молочнокислі бактерії перестають викликати скисання молока після тривалого культивування їх на середовищах без вуглеводів. Пігментні бактерії утворюють безбарвні раси при відсутності солей магнію і під впливом високої температури (Bacterium prodigiosum). Старі лабораторні розведення холерного вібріону часто перестають розріджувати желатин. Крім того, окремі бактерії дають нові різновиди та раси, значно різні між собою.
Систематика і класифікація мікроорганізмів в основному виходить із принципів, прийнятих для тварин та рослин.
Основна таксономічна одиниця в системах класифікації – вид.
Вид – група або сукупність близьких між собою організмів, які мають загальне коріння походження, характеризується визначеними морфологічними, біохімічними та фізіологічними ознаками та пристосовані до певного середовища існування.
Для встановлення виду мікробу використовують в основному наступні показники: морфологічні – форма та розміри клітини, спороутворення, рухливість, відношення до різних барвників, особливо до фарб по Граму; культуральні – характер росту на різноманітних живильних середовищах, форми колоній на щільних живильних середовищах, утворення пігменту; фізіологічні – відношення мікробів до різноманітних живильних речовин, до джерел вуглеводів та азоту, саварів та молекулярного кисню.
У мікробіології часто зустрічаються терміни «різновид »,« раса »,« штам »і« клон ». Різновидом називають певну стадію формування нових видів. Раса - термін, що позначає відокремлену в екологічному, а іноді і в морфологічному відношенні групу організмів всередині виду чи підвиду. Штамом називають чисту культуру одного і того ж виду, що відрізняється по ряду властивостей і ознак від основного виду. Клон означає популяцію (культуру), що є нащадками однієї клітини.
За міжнародними правилами, види мікроорганізмів позначаються двома словами латинською мовою. Перше слово означає рід і пишеться з великої літери, друге - видову назву і починається з малої літери.
Наприклад, Bacillus subtilis - сінна паличка, Esche richia coli - кишкова паличка.
Споріднені види бактерій з однаковими морфологічними, фізіологічними, біохімічними та іншими біологічними властивостями систематики об'єднують в роди, роди в сімейства, а останні в порядки і класи.
В даний час розроблено багато схем класифікації бактерій, але жодна з них не може вважатися цілком задовільною. Широке поширення отримала схема класифікації американського суспільства мікробіологів під редакцією А.Берджі - «Визначник бактерій». За цією схемою все мікроорганізми відносять до рослинного царства (Protophyta) і ділять на три класи: хлорофільні - синьо-зелені водорості (Schizophyceae), безхлорофільні (Schizomyceае), рикетсії і віруси (Microtatobiotes). У Радянському Союзі користуються також «Визначником бактерій і актиноміцетів» Н. А. Красильникова, який всі найпростіші рослинні організми (протофіти) ділить на дві групи - хлорофільні (Schizophyceae) і безхлорофільні (Schizomyceae). Група безхлорофільних мікроорганізмів, в свою чергу, ділиться на чотири класи: Actinomycetes, Eubacteriae, Myxobacteriae, Spirochaetae.
Нижче наводиться для прикладу принцип класифікації тільки класу бактерій.
Клас бактерій (Eubacteriae) складається з чотирьох порядків, які поділяються на сімейства, а сімейства на роди.
Порядок Eubacteriales. У нього входять одноклітинні організми кулястої, паличкоподібної і звивистою форм. Найбільш, важливі сімейства цього порядку наступні.
Сімейство Bacteriaceae - паличкоподібні грамнегативні, не утворюють спор рухливі бактерії. Це сімейство включає наступні роди:
Bacterium - Бактерії широко поширені в ґрунті, воді, повітрі, кишечнику тварин і людини, пігмент не утворюють - безкольорові;
Chromobacterium - Паличкоподібні бактерії, які відрізняються від першого роду забарвленням культур. Вони можуть бути червоного, оранжевого, жовтого, зеленого і синього кольорів;
Azotobacter - вільноіснуючі азотфіксатори;
Photobacterium - Бактерії, які на поживних середовищах виділяють фосфоричне світло;
Thiobacterium - Паличкоподібні бактерії, на мінеральних середовищах окислюють з'єднання сірки до сірчаної кислоти.
Сімейство Bacillaceae. Включає два роди: Bacillus (Рухливі палички, утворюють спори, аероби, грампозитивні) і Clostridium (Анаеробні клітини, які при спороутворенні набувають форму барабанної палички або лимон Таким чином нуклеоїд- є носієм спадкових властивостей клітини і контролює синтез специфічних білків овидну).
Сімейство Pseudomonadaceae. У нього входять бактерії що не утворюють спор, паличкоподібні грамнегативні рухомі бактерії. За фізіологічними та біохімічними ознаками ці бактерії поділені на окремі роди.
Рід Pseudomonas включає бактерії, що утворюють синій або зеленувато-синій пігмент. Мікроорганізми роду Pseudomonas denitrificans відновлюють нітрати і нітрити до вільного азоту. Бактерії роду Nitrosomonas окислюють аміак до азотистої кислоти, а роду Rhizobium утворюють бульби на коренях бобових рослин.
В порядок Eubacteriales входять бактерії з шароподібною формою клітини з сімейства Planococcaceae і зі спірально вигнутими клітинами з сімейства Spirillaceae та ін..
Порядок Ferribacteriales. Включає сімейство Ferribacteriaceae, яке об'єднує чотири роди. У цю групу включені одноклітинні кокові і паличкоподібні залізобактерії, що окислюють закис заліза.
Порядок Chlamydobacteriales. Містить ниткоподібні організми з перегородками, рухливі і нерухомі. Вони мешкають переважно в прісних водах. Всі бактерії безспорові, аероби, автотрофні або гетеротрофні. До сімейства Chlamydobacteriaceae відносяться роди Spherotilus, Leptothrix та ін.
Порядок Thiobacteriales. Група сіробактерій, куди входять одноклітинні організми кулястої, паличкоподібної і веретеновидної форми. Розміри клітин значно більші ніж у звичайних бактерій. Об'єднуються вони за здатністю окислювати сірководень в сірку, а потім в сірчану кислоту. Порядок має кілька сімейств і родів (роди Achromatium, Thiophysa та ін.)
Контрольні питання
1. Визначення мікробіології в тому числі технічної мікробіології як науки.
2. Характеристика морфологічних ознак мікроорганізмів різних систематичних груп.
3. Методи визначення розмірів бактерій.
4. Субмікроскопічна будова прокаріотичноі клітини.
5 Типи руху у мікробів.
6 Спори і спороутворення у бактерій.
7. Характеристика способів розмноження у бактерій.
8. Основні принципи класифікації бактерій.
9. Особливості будови клітинної стінки у бактерій.
10. Морфологія та класифікація актиноміцетів і грибів.
11. Особливості розмноження нижчих та вищих грибів.
12. Будова та розмноження дріжджової клітини.
14. Водорості, будова, систематика та роль у підвищенні родючості ґрунту.
15. Характерні особливості будови та розмноження вірусів.
РОЗДІЛ 2